Наращивание височной зоны: Частичное наращивание волос
Височное наращивание волос — 73 фото
Нарощенные волосы
Нарощенные волосы средней длины
Красивые нарощенные волосы
Нарощенные волосы
Наращивание волос спереди
Наращивание волос Серпухов
Наращивание волос для густоты
Скрытое наращивание для брюнеток
Наращивание челки
Микробелларго наращивание волос
Длинные волосы наращивание
Височное наращивание волос черные
Наращивание волос до и после
Загущение волос наращиванием
Капсульное наращивание волос
Наращивание волос
Височное наращивание волос
Наращивание волос на длинные волосы
Капсульное наращивание волос
Нарощенные волосы средней длины
Наращивание волос
Капсулы для наращивания волос
Каркасное наращивание волос афронаращивание
Прически для нарощенных волос
Наращивание волос в Воронеже салоны
Красиво нарощенные волосы
Наращивание волос макет
Микрокапсульное наращивание волос загущение височной части
Нарощенные пряди волос
Афронаращивание волос
Нарощенные волосы с челкой
Система наращивания волос
Ава с девочкой из фильма нарощенные волосы 2007
Наращивание волос 2021
Нарощенные волосы
Наращивание волос Сток
Капсульное афронаращивание
Частичное наращивание волос
Наращивание волос средней длины
Мастер по наращиванию волос
Капсульное наращивание волос до и после
Противопоказания к наращиванию волос
Наращивание волос загущение височной зоны
Капсульное наращивание волос Тула
Красивое наращивание волос
Наращивание челки Москва
Каркасное наращивание волос афронаращивание
Капсульное наращивание волос
Чемпионат по наращиванию волос
Микробелларго наращивание волос
Наращивание волос реклама
Нарощенные волосы
Капсульные волосы для наращивания блонд
Расчесывание блондинка волос
Зональное наращивание волос
Нарощенные волосы
Нарощенные волосы
Капсульное наращивание 75см
Загущение волос наращиванием
Наращивание волос на пышные волосы
Наращивание волос 100см Бишкек
Нарощенные пряди волос
Волосы на трессах до и после
Наращивание волос разного цвета
Наращивание волос
Наращивание височной зоны на длинные волосы
Длинные нарощенные волосы до и после
Наращивание волос по технологии FIXHAIR
Наращивание волос операция
Наращивание волос красиво
Наращивание волос объем от корней
Нарощенные черные волосы
Височное наращивание волос
Добавить
Комментарии (0)
Информация
Посетители, находящиеся в группе Гости, не могут оставлять комментарии к данной публикации.
Наращивание волос на удлиненное каре
Наращивание удлиненного каре
Наращивание височной зоны для каре
Нарощенные волосы удлиненное каре
Наращивание волос загущение височной зоны
Наращивание волос на каре
Наращивание височной зоны для каре
Наращивание волос на короткие волосы
Нарощенные черные волосы на каре
Наращивание волос височной зоны для каре
Капсульное наращивание на каре
Наращивание волос височной зоны для каре
Наращивание волос височной зоны
Наращивание волос височной зоны для каре
Наращивание волос на каре
Загущение височной зоны каре
Каре на удлинение черные волосы
Наращивание волос до каре
Красивое наращивание волос
Чёрные волосы выпрямленные каре
Наращивание каре на удлинение
Наращивание прядей на короткие волосы
Стрижка удлинениекаре до и после
Кератиновое выпрямление на Боб каре
Загущение волос височной зоны
Наращивание волос для густоты
Наращивание волос на короткую стрижку
Черное каре с белыми прядями
Нарощенные волосы
Наращивание волос для объема каре
Стрижка ровный срез на средние волосы
Каре до и после
Красивые девушки с каре брюнетки
Удлиненное каре на нарощенных волосах
Афронаращивание волос
Стрижка удлиненный Боб до и после
Каре с удлиненными передними прядями с челкой на темные волосы
Удлиненное каре на светлые волосы
Виктория Романец волосы
Капсульное наращивание
Красивые нарощенные волосы
Удлиненное каре с челкой черные волосы
Удлиненное каре без челки на длинные волосы
Капсульное ультразвуковое наращивание волос
Длинные нарощенные волосы
Каскад на нарощенные волосы
Удлинение волос без наращивания
Наращивание волос на каре
Кератиновое выпрямление на каре с челкой
Наращивание передних прядей
Прямой срез на черных волосах
Подстричь волосы под каре
Наращивание волос прически
Стрижки после каре на средние волосы
Боб с удлинением на длинные волосы
Наращивание на короткие волосы до и после
Наращивание 60 см на короткие волосы
Нарощенные волосы
Нарощенные волосы
Стрижка эффект нарощенных
Стрижка на нарощенные волосы
Удлиненное каре после длинных волос
Наращивание волос
Нарощенные волосы
Наращивание волос
Асиммитричное каре ЬОБ С У
250 Прядей микрокапсульного наращивания
Удлиненный Боб филированный
Каре-Боб с удлиненными прядями с челкой
Лонг Боб стрижка
Каре с макушкой на удлинение
Пространственно-временное расширение в месте возникновения кортикальных кальретининовых нейронов у приматов
Обзор
. 2014 26 июня; 8:50.
2014 26 июня; 8:50.
doi: 10.3389/fnana.2014.00050.
Электронная коллекция 2014.
Анна Хладник
1
, Домагой Джая
2
, Саня Дармополь
3
, Наташа Йованов-Милошевич
2
, Здравко Петанек
3
Принадлежности
- 1 Кафедра анатомии и клинической анатомии Медицинского факультета Загребского университета Загреб, Хорватия.
- 2 Кафедра неврологии, Хорватский институт исследований мозга, Медицинский факультет Загребского университета Загреб, Хорватия.

- 3 Кафедра анатомии и клинической анатомии Медицинского факультета Загребского университета Загреб, Хорватия; Кафедра неврологии, Хорватский институт исследований мозга, Медицинский факультет, Загребский университет Загреб, Хорватия.

PMID:
25018702
PMCID:
PMC4072090
DOI:
10.3389/fnana.2014.00050
Бесплатная статья ЧВК
Обзор
Ana Hladnik et al.
Фронт Нейроанат.
.
Бесплатная статья ЧВК
. 2014 26 июня; 8:50.
2014 26 июня; 8:50.
дои: 10.3389/fnana.2014.00050.
Электронная коллекция 2014.
Авторы
Анна Хладник
1
, Домагой Джая
2
, Саня Дармополь
3
, Наташа Йованов-Милошевич
2
, Здравко Петанек
3
Принадлежности
- 1 Кафедра анатомии и клинической анатомии Медицинского факультета Загребского университета Загреб, Хорватия.
- 2 Кафедра неврологии, Хорватский институт исследований мозга, Медицинский факультет Загребского университета Загреб, Хорватия.
- 3 Кафедра анатомии и клинической анатомии Медицинского факультета Загребского университета Загреб, Хорватия; Кафедра неврологии, Хорватский институт исследований мозга, Медицинский факультет, Загребский университет Загреб, Хорватия.

PMID:
25018702
PMCID:
PMC4072090
DOI:
10.3389/fnana.2014.00050
Абстрактный
Подавляющее большинство кортикальных ГАМКергических нейронов можно определить по экспрессии парвальбумина, соматостатина или кальретинина. У большинства млекопитающих интернейроны парвальбумина и соматостатина имеют постоянные пропорции, каждая из которых составляет 5-7% от общего числа нейронов. Напротив, наблюдается трехкратное увеличение доли кальретининовых интернейронов, которые не превышают 4% у грызунов и достигают 12% в областях коры головного мозга высших порядков приматов. У грызунов почти все парвальбуминовые и соматостатиновые интернейроны происходят из медиальной части субпаллиальной пролиферативной структуры, ганглиозного возвышения (ГЭ), тогда как почти все кальретининовые интернейроны происходят из его каудальной части. Пространственный паттерн происхождения кортикальных ГАМКергических нейронов из ГЭ сохраняется в мозге обезьяны и человека. Однако можно ожидать, что эволюция изменяет правила развития, чтобы сделать возможным значительное расширение популяции кальретининовых интернейронов. В течение раннего фетального периода у приматов кортикальные ГАМКергические нейроны почти полностью генерируются в субпаллиуме, как и у грызунов. Уже в это время каудальное ганглиозное возвышение (CGE) приматов показывает относительное увеличение размера и продукции кальретининовых интернейронов.
Напротив, наблюдается трехкратное увеличение доли кальретининовых интернейронов, которые не превышают 4% у грызунов и достигают 12% в областях коры головного мозга высших порядков приматов. У грызунов почти все парвальбуминовые и соматостатиновые интернейроны происходят из медиальной части субпаллиальной пролиферативной структуры, ганглиозного возвышения (ГЭ), тогда как почти все кальретининовые интернейроны происходят из его каудальной части. Пространственный паттерн происхождения кортикальных ГАМКергических нейронов из ГЭ сохраняется в мозге обезьяны и человека. Однако можно ожидать, что эволюция изменяет правила развития, чтобы сделать возможным значительное расширение популяции кальретининовых интернейронов. В течение раннего фетального периода у приматов кортикальные ГАМКергические нейроны почти полностью генерируются в субпаллиуме, как и у грызунов. Уже в это время каудальное ганглиозное возвышение (CGE) приматов показывает относительное увеличение размера и продукции кальретининовых интернейронов. Во втором триместре беременности, который является основной нейрогенетической стадией у приматов без четких коррелятов, обнаруженных у грызунов, наблюдается паллиальная продукция кортикальных ГАМКергических нейронов вместе с длительной персистенцией ГЭ. Мы предполагаем, что CGE может быть основным источником кальретининовых интернейронов для задних и латеральных областей коры, но не для лобной коры. Ассоциативная зернистая лобная кора представляет около одной трети поверхности коры и содержит почти половину корковых интернейронов кальретинина. Большинство кальретининовых интернейронов, предназначенных для лобной коры, могут генерироваться в паллиуме, особенно во вновь развившейся внешней субвентрикулярной зоне, которая становится основным пулом корковых предшественников.
Во втором триместре беременности, который является основной нейрогенетической стадией у приматов без четких коррелятов, обнаруженных у грызунов, наблюдается паллиальная продукция кортикальных ГАМКергических нейронов вместе с длительной персистенцией ГЭ. Мы предполагаем, что CGE может быть основным источником кальретининовых интернейронов для задних и латеральных областей коры, но не для лобной коры. Ассоциативная зернистая лобная кора представляет около одной трети поверхности коры и содержит почти половину корковых интернейронов кальретинина. Большинство кальретининовых интернейронов, предназначенных для лобной коры, могут генерироваться в паллиуме, особенно во вновь развившейся внешней субвентрикулярной зоне, которая становится основным пулом корковых предшественников.
Ключевые слова:
ГАМК; кальретинин; эпилепсия; ганглиозное возвышение; интернейроны; вентрикулярная зона.
Цифры
РИСУНОК 1
Вклад главной коры…
РИСУНОК 1
Вклад основных субпопуляций ГАМКергических нейронов коры в общее количество ГАМКергических…
РИСУНОК 1
Вклад основных субпопуляций ГАМКергических нейронов коры в общее число ГАМКергических нейронов у грызунов и их вклад в общее число нейронов у грызунов и приматов. (A) Демонстрирует вклад различных субпопуляций ГАМКергических нейронов у грызунов, показывая, что парвальбумин, соматостатин и кальретинин составляют более 90% всех ГАМКергических нейронов. (Б) Демонстрирует 50% увеличение вклада кортикальных ГАМКергических нейронов от грызунов (≈16%) к приматам (≈24%), что можно объяснить трехкратным увеличением числа кальретининовых нейронов (показаны красным; от ≈4% у грызунов до ≈12% от общего числа нейронов у приматов).
(A) Демонстрирует вклад различных субпопуляций ГАМКергических нейронов у грызунов, показывая, что парвальбумин, соматостатин и кальретинин составляют более 90% всех ГАМКергических нейронов. (Б) Демонстрирует 50% увеличение вклада кортикальных ГАМКергических нейронов от грызунов (≈16%) к приматам (≈24%), что можно объяснить трехкратным увеличением числа кальретининовых нейронов (показаны красным; от ≈4% у грызунов до ≈12% от общего числа нейронов у приматов).
РИСУНОК 2
Мозг зрелого человека и эмбриона…
РИСУНОК 2
Мозг зрелого человека и плода в середине беременности, демонстрирующий, что мультимодальные области более высокого порядка…
ФИГУРА 2
Мозг зрелого человека и плода примерно в середине беременности, показывающий, что мультимодальные области более высокого порядка охватывают лобную зернистую кору и большую часть теменной доли (за исключением постцентральной извилины). (А) В головном мозге человека лобная зернистая кора занимает 80% лобной доли без четкой корреляции у мышей. У человека эта кора составляет почти треть (красный цвет) всей поверхности коры, а также 80% мультимодальных ассоциативных областей, включая ассоциативные области теменной коры (синий цвет). В ассоциативных областях более высокого порядка, расположенных в лобной (красный) и теменной доле (синий), нейроны, экспрессирующие кальретинин, составляют почти 50% популяции ГАМКергических нейронов и более 10% общей популяции нейронов. В связи с предположительно меньшей долей кальретининовых нейронов в височной и затылочной коре (Ma et al., 2013), а также с учетом размеров лобной зернистой коры мы предположили, что почти половина корковых кальретининовых нейронов расположена в лобной доле. (B) Пропорции размера ассоциативной коры более высокого порядка уже установлены примерно в середине беременности. В недавнем исследовании Ma et al. (2013) пришли к выводу, что подавляющее большинство кальретининовых нейронов у человека происходит из каудального ганглиозного возвышения (КГЭ, пунктирная линия показывает контур его положения) и что в паллиальных пролиферативных зонах нет обильной продукции ГАМКергических нейронов, как предполагалось ранее.
(А) В головном мозге человека лобная зернистая кора занимает 80% лобной доли без четкой корреляции у мышей. У человека эта кора составляет почти треть (красный цвет) всей поверхности коры, а также 80% мультимодальных ассоциативных областей, включая ассоциативные области теменной коры (синий цвет). В ассоциативных областях более высокого порядка, расположенных в лобной (красный) и теменной доле (синий), нейроны, экспрессирующие кальретинин, составляют почти 50% популяции ГАМКергических нейронов и более 10% общей популяции нейронов. В связи с предположительно меньшей долей кальретининовых нейронов в височной и затылочной коре (Ma et al., 2013), а также с учетом размеров лобной зернистой коры мы предположили, что почти половина корковых кальретининовых нейронов расположена в лобной доле. (B) Пропорции размера ассоциативной коры более высокого порядка уже установлены примерно в середине беременности. В недавнем исследовании Ma et al. (2013) пришли к выводу, что подавляющее большинство кальретининовых нейронов у человека происходит из каудального ганглиозного возвышения (КГЭ, пунктирная линия показывает контур его положения) и что в паллиальных пролиферативных зонах нет обильной продукции ГАМКергических нейронов, как предполагалось ранее. (Летиник и др., 2002). Анатомические отношения между расположенной впереди префронтальной корой (составляющей почти половину кортикальных кальретининовых нейронов) и заднелатерально расположенной КГЭ (как основным источником кальретининовых нейронов) не согласуются с этим предположением (см.0119 Рисунок 5B ). Мы предполагаем, что презумпция Ma et al. (2013) могут быть верны у человека для всех областей коры только на ранней стадии плода, а затем для всех долей, за исключением лобной коры (Al-Jaberi et al., 2013).
(Летиник и др., 2002). Анатомические отношения между расположенной впереди префронтальной корой (составляющей почти половину кортикальных кальретининовых нейронов) и заднелатерально расположенной КГЭ (как основным источником кальретининовых нейронов) не согласуются с этим предположением (см.0119 Рисунок 5B ). Мы предполагаем, что презумпция Ma et al. (2013) могут быть верны у человека для всех областей коры только на ранней стадии плода, а затем для всех долей, за исключением лобной коры (Al-Jaberi et al., 2013).
РИСУНОК 3
Фронтальный срез через переднюю…
РИСУНОК 3
Фронтальный срез передней части головного мозга плода человека и мыши…
РИСУНОК 3
Фронтальный срез передней части головного мозга плода человека и мыши на ранних стадиях коркового нейрогенеза . У приматов в течение раннего фетального периода и у грызунов в течение всего нейрогенеза подавляющее большинство ГАМКергических нейронов кортикальной пластинки (КП) продуцируются в ганглиозном возвышении (ГЭ) и мигрируют тангенциально в кору головного мозга (красная пунктирная линия). Обратите внимание, что нейроны, тангенциально мигрирующие в самые отдаленные позиции в мозге мыши, могут проходить то же расстояние, что и определенные радиально мигрирующие основные глутаматергические клетки в мозге человека (желтая пунктирная линия), которые происходят в паллиальной вентрикулярной и субвентрикулярной зоне (VZ). Во время среднего триместра беременности, когда рождается большинство корковых нейронов, расхождения в масштабировании между человеком и мышью гораздо более выражены. Модифицировано из Petanjek et al. (2009 г.б).
У приматов в течение раннего фетального периода и у грызунов в течение всего нейрогенеза подавляющее большинство ГАМКергических нейронов кортикальной пластинки (КП) продуцируются в ганглиозном возвышении (ГЭ) и мигрируют тангенциально в кору головного мозга (красная пунктирная линия). Обратите внимание, что нейроны, тангенциально мигрирующие в самые отдаленные позиции в мозге мыши, могут проходить то же расстояние, что и определенные радиально мигрирующие основные глутаматергические клетки в мозге человека (желтая пунктирная линия), которые происходят в паллиальной вентрикулярной и субвентрикулярной зоне (VZ). Во время среднего триместра беременности, когда рождается большинство корковых нейронов, расхождения в масштабировании между человеком и мышью гораздо более выражены. Модифицировано из Petanjek et al. (2009 г.б).
РИСУНОК 4
Пропорции размеров взрослых грызунов…
РИСУНОК 4
Соотношение размеров мозга зрелых грызунов и приматов, а также развивающегося…
РИСУНОК 4
Соотношение размеров мозга зрелых грызунов и приматов, а также мозга развивающегося и зрелого человека . Вид сверху на взрослую мышь (A) , макаку-резус (A) и человеческий мозг (B) , а также мозг человеческого плода в середине беременности (A) и в срок (B) . (A) Размер головного мозга плода человека уже в середине беременности достигает размера мозга взрослой макаки-резус. Тем не менее, мозг взрослой макаки-резус почти в 100 раз больше, чем мозг взрослой мыши. (B) Мозг взрослого человека примерно в 3–4 раза больше, чем мозг новорожденного, размер которого достигает размера мозга взрослого шимпанзе. Обратите внимание, что картина гирификации в мозге новорожденного человека близка к той, которая наблюдается у взрослых. Впускное отверстие в середине рисунка представляет собой интегративное фото всех мозгов, показанных на 9-й странице.0119 A и B , которые демонстрируют их реальные пропорции (правый мозг в верхнем ряду вставки от плода в начале третьего триместра беременности).
Вид сверху на взрослую мышь (A) , макаку-резус (A) и человеческий мозг (B) , а также мозг человеческого плода в середине беременности (A) и в срок (B) . (A) Размер головного мозга плода человека уже в середине беременности достигает размера мозга взрослой макаки-резус. Тем не менее, мозг взрослой макаки-резус почти в 100 раз больше, чем мозг взрослой мыши. (B) Мозг взрослого человека примерно в 3–4 раза больше, чем мозг новорожденного, размер которого достигает размера мозга взрослого шимпанзе. Обратите внимание, что картина гирификации в мозге новорожденного человека близка к той, которая наблюдается у взрослых. Впускное отверстие в середине рисунка представляет собой интегративное фото всех мозгов, показанных на 9-й странице.0119 A и B , которые демонстрируют их реальные пропорции (правый мозг в верхнем ряду вставки от плода в начале третьего триместра беременности).
РИСУНОК 5
Места происхождения различных…
РИСУНОК 5
Места происхождения различных популяций ГАМКергических нейронов . Срезы, окрашенные по Нисслю: (A)…
Срезы, окрашенные по Нисслю: (A)…
РИСУНОК 5
Места происхождения различных популяций ГАМКергических нейронов . Срезы, окрашенные по Нисслю: (А) фронтальный срез на уровне таламуса и (В) горизонтальный срез тела мозолистого тела (СС) в конце среднего триместра беременности. (A) На срезе показан основной пролиферативный компартмент мантийного мозга человека, внешняя субвентрикулярная зона (oSVZ), которая является наиболее вероятным кандидатом на роль основного источника дополнительного пула предшественников ГАМКергических нейронов (Petanjek and Kostovic, 2012). В пользу этой точки зрения говорит тот факт, что уже в ранний фетальный период клетки-предшественники коры головного мозга человека в лобной доле обладают способностью генерировать тормозные интернейроны (Al-Jaberi et al., 2013). (B) У мышей подавляющее большинство экспрессирующих парвальбумин и соматостатин (черная линия) ГАМКергических нейронов корковой пластинки (CP) происходят из медиального ганглиозного возвышения (mGE), в то время как экспрессирующие кальретинин происходят из каудального ганглиозного возвышения (CGE). Прогениторы из латерального ганглиозного возвышения (lGE) дают ГАМКергические нейроны в основном для подкорковых структур. Этот пространственный паттерн сохраняется в течение раннего триместра беременности у приматов, но с экспоненциальным увеличением пула предшественников cGE (Hansen et al., 2013). Мы предполагаем, что во время среднего триместра беременности, когда продуцируется большинство кортикальных нейронов, cGE остается основным источником кортикальных нейронов кальретинина (синяя пунктирная линия) для теменной, затылочной и височной долей (заднелатеральный конечный мозг), тогда как лобная доля (красная пунктирная линия) снабжается кальретининовыми нейронами из oSVZ (см. Рисунок 2B ). Не только анатомические отношения (довольно удаленное положение лобной доли от ХГЭ), не согласующиеся с исключительной продукцией кальретининовых нейронов из ХГЭ (Ma et al., 2013), но и трехкратное увеличение доли кальретининовых нейронов в лобной коре не может быть достигнуто за счет увеличения пула и длительного производства предшественников в cGE.
Прогениторы из латерального ганглиозного возвышения (lGE) дают ГАМКергические нейроны в основном для подкорковых структур. Этот пространственный паттерн сохраняется в течение раннего триместра беременности у приматов, но с экспоненциальным увеличением пула предшественников cGE (Hansen et al., 2013). Мы предполагаем, что во время среднего триместра беременности, когда продуцируется большинство кортикальных нейронов, cGE остается основным источником кортикальных нейронов кальретинина (синяя пунктирная линия) для теменной, затылочной и височной долей (заднелатеральный конечный мозг), тогда как лобная доля (красная пунктирная линия) снабжается кальретининовыми нейронами из oSVZ (см. Рисунок 2B ). Не только анатомические отношения (довольно удаленное положение лобной доли от ХГЭ), не согласующиеся с исключительной продукцией кальретининовых нейронов из ХГЭ (Ma et al., 2013), но и трехкратное увеличение доли кальретининовых нейронов в лобной коре не может быть достигнуто за счет увеличения пула и длительного производства предшественников в cGE.
См. это изображение и информацию об авторских правах в PMC
Похожие статьи
Специфическое для приматов происхождение и миграция кортикальных ГАМКергических нейронов.
Петанек З., Костович И., Эсклапез М.
Петанжек З. и др.
Фронт Нейроанат. 2009 27 ноября; 3:26. doi: 10.3389/нейро.05.026.2009. Электронная коллекция 2009.
Фронт Нейроанат. 2009.PMID: 20011218
Бесплатная статья ЧВК.Потеря COUP-TFI изменяет баланс между кортикальными интернейронами, происходящими из каудального ганглиозного возвышения и медиального ганглиозного возвышения, и приводит к резистентности к эпилепсии.
Лодато С., Томасси Г.С., Де Леонибус Э., Узкатеги Ю.Г., Андольфи Г., Арментано М., Тузот А., Газтелу Дж.
М., Арлотта П., Менендес де ла Прида Л., Студер М.
Лодато С. и соавт.
Дж. Нейроски. 2011 23 марта; 31 (12): 4650-62. doi: 10.1523/JNEUROSCI.6580-10.2011.
Дж. Нейроски. 2011.PMID: 21430164
Бесплатная статья ЧВК.Сложность экспрессирующих кальретинин предшественников в коре головного мозга человека.
Радонич Н.В., Ортега Ю.А., Меми Ф., Дионн К., Яковцевски И., Зечевич Н.
Радонич Н.В. и соавт.
Фронт Нейроанат. 2014 13 августа; 8:82. doi: 10.3389/fnana.2014.00082. Электронная коллекция 2014.
Фронт Нейроанат. 2014.PMID: 25165435
Бесплатная статья ЧВК.Перспективы развития коркового интернейронного разнообразия.
Фишелл Г.
Фишел Г.
Novartis обнаружила Symp. 2007; 288:21-35; обсуждение 35-44, 96-8.
Novartis обнаружила Symp. 2007.PMID: 18494250
Обзор.
Нейроны кальретинина неокортекса у приматов: увеличение пропорций и структура микросхемы.
Джая Д., Хладник А., Бичанич И., Бакович М., Петанек З.
Джая Д и др.
Фронт Нейроанат. 2014 25 сент.; 8:103. doi: 10.3389/fnana.2014.00103. Электронная коллекция 2014.
Фронт Нейроанат. 2014.PMID: 25309344
Бесплатная статья ЧВК.Обзор.
 М., Арлотта П., Менендес де ла Прида Л., Студер М.
М., Арлотта П., Менендес де ла Прида Л., Студер М.
Посмотреть все похожие статьи
Цитируется
У мышей с дефицитом PlexinA1 наблюдается снижение плотности клеток и повышенный окислительный стресс в интернейронах, экспрессирующих парвальбумин, в медиальной префронтальной коре.
Джахан М.С., Цузуки Т., Ито Т.
, Бхуйян М.Э., Такахаши И., Такамацу Х., Куманогох А., Негиши Т., Юкава К.
Джахан М.С. и др.
IBRO Neurosci Rep. 2022 12 ноября; 13:500-512. doi: 10.1016/j.ibneur.2022.11.002. электронная коллекция 2022 дек.
Представитель IBRO Neurosci, 2022 г.PMID: 36451778
Бесплатная статья ЧВК.Нарушение сети верхнего коркового слоя при шизофрении.
Батюк М.Ю., Тайлер Т., Драгичевич К., Мей С., Ридбирк Р., Петухов В., Девятияров Р., Седмак Д., Франк Э., Фехер В., Хабек Н., Ху К., Иголкина А., Росзик Л., Пфистерер У., Гарсия-Гонсалес Д, Петанжек З., Адоржан И., Харченко П.В., Ходосевич К.
Батюк М.Ю. и соавт.
Научная реклама 2022 14 окт;8(41):eabn8367. doi: 10.1126/sciadv.abn8367. Epub 2022 12 октября.
Научная реклама 2022.PMID: 36223459Бесплатная статья ЧВК.
Серотонинергическая регуляция выживания биполярных клеток в развивающейся коре головного мозга.
Вонг Ф.К., Селтен М., Росес-Новелла С., Шринивасан В., Паллас-Базарра Н., Серафеймиду-Пулиу Э., Хануш-Годой А., Оозир Ф., Эдвардс Р., Марин О.
Вонг Ф.К. и др.
Cell Rep. 5 июля 2022 г .; 40 (1): 111037. doi: 10.1016/j.celrep.2022.111037.
Представитель ячейки 2022.PMID: 35793629
Бесплатная статья ЧВК.Отличительные характеристики соматостатиновых нейронов в человеческом мозге.
Бановац И., Седмак Д., Эсклапес М., Петанжек З.
Бановац I и др.
Мол Нейробиол. 2022 авг; 59 (8): 4953-4965. doi: 10.1007/s12035-022-02892-6. Epub 2022 4 июня.
Мол Нейробиол. 2022.PMID: 35665897
Измененный паттерн предшественников интернейронов трисомии 21.
Гиффин-Рао Ю., Шэн Дж., Стрэнд Б., Сюй К.
, Хуан Л., Медо М., Рисгаард К.А., Дантинн С., Мохан С., Кешан А., Дейли Р.А. младший, Левеск Б., Амундсон Л., Риз Р., Соуза АММ, Тао И, Ван Д, Чжан С.К., Бхаттачарья А.
Гиффин-Рао Ю. и др.
Отчеты о стволовых клетках. 2022 14 июня; 17 (6): 1366-1379. doi: 10.1016/j.stemcr.2022.05.001. Epub 2022 26 мая.
Отчеты о стволовых клетках. 2022.PMID: 35623352
Бесплатная статья ЧВК.
 , Бхуйян М.Э., Такахаши И., Такамацу Х., Куманогох А., Негиши Т., Юкава К.
, Бхуйян М.Э., Такахаши И., Такамацу Х., Куманогох А., Негиши Т., Юкава К.
 , Хуан Л., Медо М., Рисгаард К.А., Дантинн С., Мохан С., Кешан А., Дейли Р.А. младший, Левеск Б., Амундсон Л., Риз Р., Соуза АММ, Тао И, Ван Д, Чжан С.К., Бхаттачарья А.
, Хуан Л., Медо М., Рисгаард К.А., Дантинн С., Мохан С., Кешан А., Дейли Р.А. младший, Левеск Б., Амундсон Л., Риз Р., Соуза АММ, Тао И, Ван Д, Чжан С.К., Бхаттачарья А.Просмотреть все статьи «Цитируется по»
Рекомендации
Аль-Джабери Н., Линдси С., Сарма С., Баятти Н., Клоури Г.Дж. (2013). Раннее эмбриональное развитие неокортикальных ГАМКергических интернейронов человека. Церебр. кора. 10.1093/cercor/bht254 [Epub перед печатью]
—
DOI
—
ЧВК
—
пабмед
Андерсон С.
А., Казновски С.Е., Хорн С., Рубинштейн Дж.Л., Макконнелл С.К. (2002). Различное происхождение проекционных нейронов и интернейронов неокортекса in vivo. Церебр. Кора 12 702–709 10.1093/cercor/12.7.702—
DOI
—
пабмед
Антипа М., Фо К., Эйхеле Г., Парнавелас Дж. Г., Эндрюс В. Д. (2011). Дифференциальная экспрессия генов в мигрирующих потоках интернейронов коры. Евро. Дж. Нейроски. 34 1584–1594 10.1111/j.1460-9568.2011.07896.x
—
DOI
—
ЧВК
—
пабмед
Асколи Г.
А., Алонсо-Нанкларес Л., Андерсон С. А., Баррионуево Г., Бенавидес-Пиччоне Р., Беркхальтер А. и др. (2008). Терминология Petilla: номенклатура особенностей ГАМКергических интернейронов коры головного мозга. Нац. Преподобный Нейроски. 9 557–568 10.1038/nrn2402—
DOI
—
ЧВК
—
пабмед
Баринка Ф., Друга Р. (2010). Экспрессия кальретинина в неокортексе млекопитающих: обзор. Физиол. Рез. 59 665–677
—
пабмед
 А., Казновски С.Е., Хорн С., Рубинштейн Дж.Л., Макконнелл С.К. (2002). Различное происхождение проекционных нейронов и интернейронов неокортекса in vivo. Церебр. Кора 12 702–709 10.1093/cercor/12.7.702
А., Казновски С.Е., Хорн С., Рубинштейн Дж.Л., Макконнелл С.К. (2002). Различное происхождение проекционных нейронов и интернейронов неокортекса in vivo. Церебр. Кора 12 702–709 10.1093/cercor/12.7.702 А., Алонсо-Нанкларес Л., Андерсон С. А., Баррионуево Г., Бенавидес-Пиччоне Р., Беркхальтер А. и др. (2008). Терминология Petilla: номенклатура особенностей ГАМКергических интернейронов коры головного мозга. Нац. Преподобный Нейроски. 9 557–568 10.1038/nrn2402
А., Алонсо-Нанкларес Л., Андерсон С. А., Баррионуево Г., Бенавидес-Пиччоне Р., Беркхальтер А. и др. (2008). Терминология Petilla: номенклатура особенностей ГАМКергических интернейронов коры головного мозга. Нац. Преподобный Нейроски. 9 557–568 10.1038/nrn2402 Типы публикаций
Что такое временные данные?—ArcMap | Документация
- Источники данных
- Структуры данных
- Форматы даты и времени
- Поле идентификатора дорожки
Временные данные можно добавить как слой в ArcMap или ArcGlobe с помощью дополнительного модуля ArcGIS Tracking Analyst. Эти данные включают информацию о времени и дате для географических местоположений, что позволяет отслеживать наблюдения в реальном времени и ранее задокументированные наблюдения. Эти наблюдения могут быть дискретными, например, удары молнии, или непрерывными, например, маршруты перевозки грузов и траектории полета.
Эти данные включают информацию о времени и дате для географических местоположений, что позволяет отслеживать наблюдения в реальном времени и ранее задокументированные наблюдения. Эти наблюдения могут быть дискретными, например, удары молнии, или непрерывными, например, маршруты перевозки грузов и траектории полета.
Существует несколько способов добавления временных данных на карту в качестве слоя отслеживания. После добавления на карту слоя отслеживания вы можете использовать расширенные возможности Tracking Analyst для анализа и визуализации ваших данных уникальными способами на основе временных значений в данных. Например, вы можете нарисовать линии трека, чтобы показать путь объекта во времени, или использовать временные окна для воспроизведения ваших данных, показывающих только подмножество данных в любой момент времени.
Узнайте больше о параметрах временных символов, доступных в Tracking Analyst
Источники данных
Tracking Analyst принимает сохраненные временные данные или данные фиксированного времени, а также данные реального времени из нескольких источников.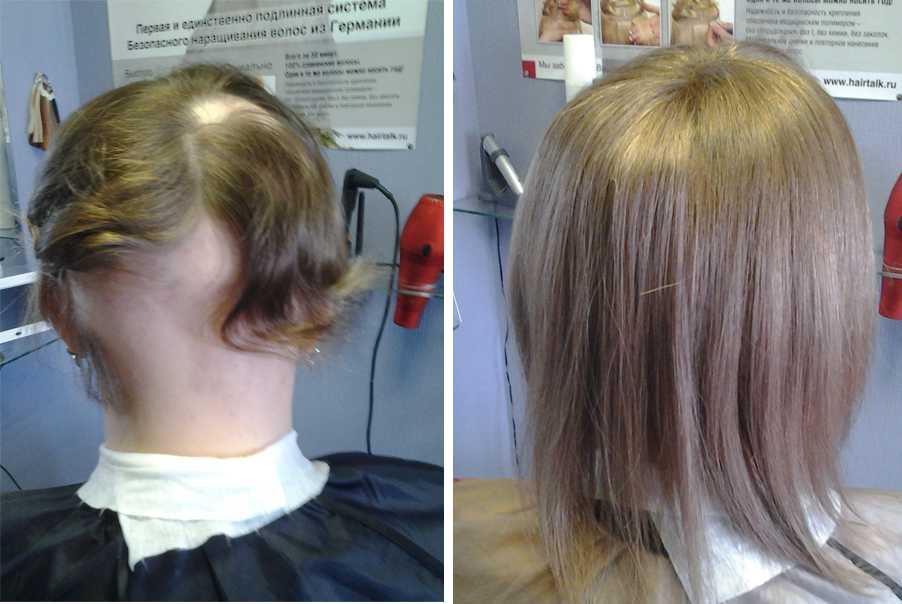
| Fixed-time sources | Real-time sources |
|---|---|
Shapefiles | Esri Tracking Server |
Geodatabase feature classes | Locally connected GPS |
Узнайте больше об источниках и структурах данных реального времени
Структуры данных
Tracking Analyst принимает следующие структуры данных как из источников реального времени, так и из источников фиксированного времени:
- Простое событие
- Сложное стационарное событие
- Сложное динамическое событие
Дополнительные сведения о простых и сложных временных событиях
Форматы даты и времени
Любые данные, используемые в Tracking Analyst, должны содержать информацию о дате и/или времени в отметке времени. Tracking Analyst поддерживает широкий спектр форматов времени и даты, которые частично зависят от форматов даты и времени в Windows. Мастер добавления временных данных позволяет выбирать разные локали и выбирать из нескольких форматов времени и даты Windows для каждой локали. Это позволяет Tracking Analyst извлекать информацию о времени и дате, содержащуюся в текстовых полях, во многих различных региональных форматах. Форматы времени и даты, хранящиеся в поле типа date, могут быть автоматически обнаружены Tracking Analyst. Если ваши данные содержат информацию о дате и времени, они должны содержаться в одном поле, чтобы их можно было использовать в Tracking Analyst.
Tracking Analyst поддерживает широкий спектр форматов времени и даты, которые частично зависят от форматов даты и времени в Windows. Мастер добавления временных данных позволяет выбирать разные локали и выбирать из нескольких форматов времени и даты Windows для каждой локали. Это позволяет Tracking Analyst извлекать информацию о времени и дате, содержащуюся в текстовых полях, во многих различных региональных форматах. Форматы времени и даты, хранящиеся в поле типа date, могут быть автоматически обнаружены Tracking Analyst. Если ваши данные содержат информацию о дате и времени, они должны содержаться в одном поле, чтобы их можно было использовать в Tracking Analyst.
Tracking Analyst поддерживает даты с 1 января 4713 г. до н.э. по 31 декабря 9999 г. н.э. Архитектура Tracking Analyst поддерживает миллисекунды, но есть некоторые ограничения на добавление данных с таким уровнем точности. Миллисекунды поддерживаются только для данных в реальном времени, а не для данных отслеживания в фиксированное время.
Поле идентификатора дорожки
Независимо от того, работаете ли вы с простыми или сложными временными событиями, концепция поля идентификатора дорожки или поля идентификатора дорожки чрезвычайно важна в Tracking Analyst. Поле Track ID не обязательно для всех слоев отслеживания, но оно может расширить возможности Tracking Analyst, если ваши данные отслеживания содержат несколько отслеживаемых объектов.
Поле Track ID должно содержать значения данных, которые однозначно идентифицируют отдельные объекты в ваших данных. Идентификатор дорожки можно использовать для группировки различных наблюдений одного и того же объекта в целях отображения и анализа. Например, если вы отслеживаете несколько грузовиков доставки, присвоение каждому грузовику уникального идентификатора отслеживания позволяет отслеживать маршрут каждого грузовика в отдельности. Существуют специальные варианты символов, использующие идентификатор дорожки. Например, линии треков позволят вам соединить позиции каждого грузовика в течение дня и воссоздать его маршрут доставки, подобно соединению точек.