Фенотипы человека: Фенотип человека: Фенотипы европеоидной расы Европы
РАСОВЫЕ ФЕНОТИПЫ ЧЕЛОВЕКА. Рекомендую к чтению. — «Правительство на другой планете живет, родной» (С) Кин — дза
- РАСОВЫЕ ФЕНОТИПЫ ЧЕЛОВЕКА. Рекомендую к чтению.
- samolet73
- April 9th, 2014
Оригинал взят у sofia_vb_888 в РАСОВЫЕ ФЕНОТИПЫ ЧЕЛОВЕКА
Впервые на русском языке публикуется перечень расовых фенотипов (более 130) иллюстрированных изображениями усреднённого представителя фенотипа, до этого на русском языке можно было найти только подборки фотографий различных фенотипов – сложно воспринимаемые и интерпретируемы за счёт разных ракурсов, разной постановки света и вкрапления не чётко выраженных и смешанных фенотипов.
— БЕЛАЯ (ЕВРОПИОЙДНАЯ) РАСА альтернативный взгляд на фенотипы Белых можно прочесть на Форуме Вольных Русских.
— НЕГРОИДНАЯ РАСА
— МОНГОЛОИДНАЯ РАСА
— АВСТРАЛОИДНАЯ РАСА
— АМЕРИКАНСКАЯ РАСАПриведённые усреднённые изображение фенотипов получены путём наложения друг на друга фотографий нескольких классических представителей типа.

Описание фенотипов и изображения взяты с англоязычного сайта: http://www.humanphenotypes.com/ . Который в настоящее время закрыт для обновления.
ОСНОВНЫЕ ОПРЕДЕЛЕНИЯ
Раса — система популяций человека, характеризующаяся сходством по комплексу определённых наследственных биологических признаков, имеющим внешнее фенотипическое проявление и сформировавшимся в определенном географическом регион (Рогинский А. А., Левин Н. Г. Антропология: Учебник для студентов ун-тов. — З-е изд., перераб.и доп. — М., Высшая школа, 1978 (с.341)).
Раса — Исторически сложившаяся группа человечества (европеоидная, негроидная, монголоидная), объединенная общностью происхождения и общностью наследственных физических признаков: цветом кожи и волос, разрезом глаз, формой черепа и т.п. (Толковый словарь Ефремовой. Т. Ф. Ефремова. 2000).
Нация (от лат. natio — племя, народ) — социально-экономическая, культурно-политическая и психо-биологическая общность, осознающая себя субъектом исторического процесса, продолжающаяся во времени за счёт генетической и культурной преемственности.
Существует два основных подхода к пониманию нации: как политической общности граждан определенного государства и как этнической общности с единым языком и самосознанием. Есть мнение, что основополагающей чертой нации является высокий уровень самоорганизации (национальное государство), в противном случае следует говорить не о нации, а о народе или этносе, хотя изначально слова «нация», «народ», «этнос», «племя» — являются синонимами (авторское определение).Русские – это Нация, обладающая устойчивыми генетически передающимися психофизиологическими признаками, национальным самосознанием и уникальной культурой. Русские являются носителями фено- и генотипа Белой Расы. Русская культура является частью индоевропейской культуры (Из программы Объединения Русские).
По мнению автора поста Русским наиболее свойственны фенотипы Нордида, Балтида, Альпенида их различных подтипов и вариаций, а так же Северный Понтид, Восточный Европид, Уралид.
Фенотип — комбинация физических человеческих черт. Средний фенотип различных мест в мире отличается. Определённые Фенотипы свойственны той или иной расе. Большинство людей в мире переставляют — смесь 2 фенотипов, у некоторых даже есть 3 вовлеченных фенотипа.
ПРИМЕЧАНИЕ
Мы привыкли гордиться своей расой и нацией, но при этом часть из нас отрицает этно-генетическую составляющую нации, а другая часть имеет смутное представление о фенологических различиях наций и рас, хотя данные знания на наш взгляд необходимы для завершения самоидентификации националиста и просто всесторонне развитой личности.
Из-за тотальной идеологии мультикультуризма развитие расологии существенно тормозится, как в России, так и за рубежом. Например в мире до сих пор не существует единой общепризнанной расовой классификации, часть классификаций объединяет различные не соприкасающиеся фенотипы в одну расу по формальным признакам. Другим классификациям свойственно выделать каждый фенотип с ответвлениями в отдельную расу.
На наш взгляд такой подход не является правильным, так как сужает расу до рамок меньше чем нация и народ, хотя более адекватным реальной ситуации является как раз включение ряда близких народов в одну расу. Кроме того ряд фенотипов редко встречается в чистом виде (но при этом смешивается с фенотипами своего или расовоблизких народов), другие фенотипы не имеют прямую генетическую передачу в отличие от расовых или национальных особенностей (передача того или иного фенотипа может чередоваться в семье через одно-два поколения по принципам описанным классическими законами наследования и изменчивости).Для составления данного материала нами была использована наиболее часто используемая классификация:
— европиойды – включает исконное население Европы и народы, граничащие с Европой, при наличии у них доминирующих европиойдный признаков (в сравнении с признаками других рас), обладающие общими устойчивыми морфологическими признаками.
— негройды – включает в себя исконное население Африки и ближайшие народны при наличии у них доминирующих негроидных признаков, обладающие общими устойчивыми морфологическими признаками.
— монголойды – исконное население Азии, Сибири и Дальнего Востока, обладающие общими устойчивыми морфологическими признаками.
— австралойды – исконное население Австралии и близлежащих островов, обладающие общими устойчивыми морфологическими признаками.
— америнды – исконное население Северной и Южной (Латинской) Америки, обладающие общими устойчивыми морфологическими признаками.
(В некоторых популярных классификациях монголойды и америнды объединены в азиатско-американскую расу, а негроиды и австралойды в — экваториальную или австрало-негроидную расу)
Мы исходим из того, что один или несколько близких фенотипов образует народ или нацию, а близкие народы образуют расу. При этом один и тот же фенотип может, встречается у разных народов, однако не может быть встречен у разных рас, за редким исключением.
Так же надо отметить, что существует ряд пограничных фенотипов, включение которых в ту или иную расу является спорным из-за наличия у них равного проявления морфологических особенностей различных рас.
Источник: ДПНИ.орг
Если Ваш провайдер блокирует ДПНИ.орг
ВЫ ВСЕ РАВНО МОЖЕТЕ СМОТРЕТЬ САЙТ:
1) через специальный Tor-браузер – пользоваться тором так же просто как и обычным браузером, просто скачайте, установите и запустите кликнув на иконку с луковицей. Скачать Tor можно бесплатно на официальном сайте https://www.torproject.org/ нажав кнопку Download Tor
2) Так же вам могут помочь анонимайзеры – специальные сайты, которые позволяют заходить на заблокированные сайты и скрывать ваш IP. Аномайзеров достаточно много и вы можете выбрать любой по вкусу — http://yandex.ua/yandsearch?text=анонимайзер&clid=46510&tld=ua&lr=145
3) Плагин для Файрфокс — самое простое решение послать систему подальше и быть в курсе правого взгляда на новости.
Tags: для_подумать, национализм, полезное, расология

 Существует два основных подхода к пониманию нации: как политической общности граждан определенного государства и как этнической общности с единым языком и самосознанием. Есть мнение, что основополагающей чертой нации является высокий уровень самоорганизации (национальное государство), в противном случае следует говорить не о нации, а о народе или этносе, хотя изначально слова «нация», «народ», «этнос», «племя» — являются синонимами (авторское определение).
Существует два основных подхода к пониманию нации: как политической общности граждан определенного государства и как этнической общности с единым языком и самосознанием. Есть мнение, что основополагающей чертой нации является высокий уровень самоорганизации (национальное государство), в противном случае следует говорить не о нации, а о народе или этносе, хотя изначально слова «нация», «народ», «этнос», «племя» — являются синонимами (авторское определение).
 На наш взгляд такой подход не является правильным, так как сужает расу до рамок меньше чем нация и народ, хотя более адекватным реальной ситуации является как раз включение ряда близких народов в одну расу. Кроме того ряд фенотипов редко встречается в чистом виде (но при этом смешивается с фенотипами своего или расовоблизких народов), другие фенотипы не имеют прямую генетическую передачу в отличие от расовых или национальных особенностей (передача того или иного фенотипа может чередоваться в семье через одно-два поколения по принципам описанным классическими законами наследования и изменчивости).
На наш взгляд такой подход не является правильным, так как сужает расу до рамок меньше чем нация и народ, хотя более адекватным реальной ситуации является как раз включение ряда близких народов в одну расу. Кроме того ряд фенотипов редко встречается в чистом виде (но при этом смешивается с фенотипами своего или расовоблизких народов), другие фенотипы не имеют прямую генетическую передачу в отличие от расовых или национальных особенностей (передача того или иного фенотипа может чередоваться в семье через одно-два поколения по принципам описанным классическими законами наследования и изменчивости).

Фенотип антигенов эритроцитов и индекс сенсибилизации у доноров компонентов крови и пациентов г.
 Астаны
Астаны
Ж.К. Буркитбаев, Р.Б. Меирбекова, Е.Н. Курганская, Л.Л. Карп, Р.З. Магзумова
Научно-производственный центр трансфузиологии, г. Астана, Казахстан
Трансфузиология №2, 2013
Резюме
В результате проведенного исследования установлено большое разнообразие фенотипов в системе Резус, а также достаточно высокая степень сенсибилизации у реципиентов и высокий индекс сенсибилизации населения города Астаны.
Ключевые слова: антигенный состав эритроцитов.
Введение
Исследование распространенности у доноров основных клинически значимых антигенов эритроцитов, определение индекса аллоиммунизации является непременным условием адекватной профилактики посттрансфузионных реакций и осложнений гемолитического типа. Определение группы крови, резус-принадлежности и типирование антигенов эритроцитов имеет большое значение при проведении гемотрансфузий для подбора совместимых пар донор-реципиент и профилактики наиболее опасных для жизни реципиентов посттрансфузионных осложнений гемолитического типа [1–16]. Методика агглютинации в геле была разработана с целью стандартизации реакций гемагглютинации и получения достоверных результатов. Тестирование на индивидуальную совместимость крови донора и реципиента по антигенам эритроцитов не заменяет обязательное иммуногематологическое исследование, а лишь дополняет его. Использование гелевой системы позволяет также снизить риск заражения персонала даже при работе с потенциально инфицированными образцами [2]. Благодаря высокой чувствительности, стандартности и хорошей воспроизводимости результатов, методы агглютинации в геле получили широкое распространение [3].
Методика агглютинации в геле была разработана с целью стандартизации реакций гемагглютинации и получения достоверных результатов. Тестирование на индивидуальную совместимость крови донора и реципиента по антигенам эритроцитов не заменяет обязательное иммуногематологическое исследование, а лишь дополняет его. Использование гелевой системы позволяет также снизить риск заражения персонала даже при работе с потенциально инфицированными образцами [2]. Благодаря высокой чувствительности, стандартности и хорошей воспроизводимости результатов, методы агглютинации в геле получили широкое распространение [3].
Цель и задачи исследования
Целью работы явилось изучение частоты встречаемости антигенов эритроцитов и определение уровня сенсибилизации населения г. Астаны.
Материал и методы исследования
Материалом исследования служили 119882 образцов крови доноров и 6313 реципиентов за период 2010–2012 гг. Возраст доноров колебался от 18 до 60 лет. Средний возраст доноров составил 43,4±5,1 года. Возраст больных колебался от 0 до 77 лет. Средний возраст больных составил 37,9±9,5 года. Среди доноров было 64736 мужчин (54,0%) и 55146 – женщин (46,0%). Среди реципиентов мужчины составили 66,9%, женщины – 33,1%. Контингент обследованных пациентов: реципиенты, в анамнезе которых многократные трансфузии; беременные, входившие в группу риска по аллоиммунизации; дети с гемолитической болезнью новорожденных. Все образцы крови доноров исследовались на групповую принадлежность по системе АВО перекрестным методом с использованием моноклональных антител, стандартных эритроцитов, а также методом микроколоночной агглютинации с использованием кассет ORTHO BioVue. Образцы крови реципиентов исследовались на групповую принадлежность по системе АВО с применением гелевой методики Scangel. А нтиген D с истемы Р езус выявляли с помощью Цоликлона АнтиD Супер (ООО «Гематолог», Москва) и с применением метода микроколоночной агглютинации с использованием кассет ORTHO BioVue, дополнительно все RH(D)-отрицательные доноры (3464 из 119 тысяч) исследовались на наличие вариантных и слабых форм антигена D с помощью моноклональных антител анти-Rh2(D)/ RhW1 в реакции Кумбса с применением гелевой методики Scangel (Карты Scangel Кумбс анти-IgG, C3d), «BioRad Laboratories», США).
Средний возраст доноров составил 43,4±5,1 года. Возраст больных колебался от 0 до 77 лет. Средний возраст больных составил 37,9±9,5 года. Среди доноров было 64736 мужчин (54,0%) и 55146 – женщин (46,0%). Среди реципиентов мужчины составили 66,9%, женщины – 33,1%. Контингент обследованных пациентов: реципиенты, в анамнезе которых многократные трансфузии; беременные, входившие в группу риска по аллоиммунизации; дети с гемолитической болезнью новорожденных. Все образцы крови доноров исследовались на групповую принадлежность по системе АВО перекрестным методом с использованием моноклональных антител, стандартных эритроцитов, а также методом микроколоночной агглютинации с использованием кассет ORTHO BioVue. Образцы крови реципиентов исследовались на групповую принадлежность по системе АВО с применением гелевой методики Scangel. А нтиген D с истемы Р езус выявляли с помощью Цоликлона АнтиD Супер (ООО «Гематолог», Москва) и с применением метода микроколоночной агглютинации с использованием кассет ORTHO BioVue, дополнительно все RH(D)-отрицательные доноры (3464 из 119 тысяч) исследовались на наличие вариантных и слабых форм антигена D с помощью моноклональных антител анти-Rh2(D)/ RhW1 в реакции Кумбса с применением гелевой методики Scangel (Карты Scangel Кумбс анти-IgG, C3d), «BioRad Laboratories», США). Все образцы крови доноров тестировались на наличие антигена К системы Кell с использованием моноклональных антител анти-К Супер (ООО «Гематолог», Москва) и с помощью метода микроколоночной агглютинации с использованием ORTHO BioVue анти-К. Все RH(D)-отрицательные доноры, а также «кадровые» RH(D)-положительные доноры были фенотипированы по антигенам С, с, Е, е системы Резус с использованием метода микроколоночной агглютинации с использованием кассет RH/K ORTHO BioVue, все реципиенты были фенотипированы по антигенам С, с, Е, е системы Резус с помощью гелевого метода с использованием карт Scangel Моноклональные RH/Kell производст- ва «Bio-Rad Laboratories», США. Во всех образцах крови доноров и реципиентов был проведен скрининг антиэритроцитарных аллоантител с помощью реакции Кумбса, выполненной с применением гелевой методики Scangel (Карты Scangel Кумбс анти-IgG, C3d), с эритроцитами Scangel I-II-III производства фирмы «Bio-Rad Laboratories», США-Франция). При положительных результатах скрининга проводилась идентификация анти- тел с эритроцитами ScanPanel из 10 линий клеток, «Bio-Rad laboratories», США.
Все образцы крови доноров тестировались на наличие антигена К системы Кell с использованием моноклональных антител анти-К Супер (ООО «Гематолог», Москва) и с помощью метода микроколоночной агглютинации с использованием ORTHO BioVue анти-К. Все RH(D)-отрицательные доноры, а также «кадровые» RH(D)-положительные доноры были фенотипированы по антигенам С, с, Е, е системы Резус с использованием метода микроколоночной агглютинации с использованием кассет RH/K ORTHO BioVue, все реципиенты были фенотипированы по антигенам С, с, Е, е системы Резус с помощью гелевого метода с использованием карт Scangel Моноклональные RH/Kell производст- ва «Bio-Rad Laboratories», США. Во всех образцах крови доноров и реципиентов был проведен скрининг антиэритроцитарных аллоантител с помощью реакции Кумбса, выполненной с применением гелевой методики Scangel (Карты Scangel Кумбс анти-IgG, C3d), с эритроцитами Scangel I-II-III производства фирмы «Bio-Rad Laboratories», США-Франция). При положительных результатах скрининга проводилась идентификация анти- тел с эритроцитами ScanPanel из 10 линий клеток, «Bio-Rad laboratories», США.
Результаты и обсуждения
Нами было проанализировано распределение групп крови системы АВО у доноров и реципиентов (таблица 1).
Полученные данные свидетельствуют о том, что распределение групп крови по системе АВО у доноров и пациентов сходны. Преобладающими являются O и А группы крови. Выявленная нами частота встречаемости антигена А2 в группе крови А и АВ составляет для доноров 0,32%, а для пациентов 3,2%. Выявленное нами соотношение Rh(D)-положительных и Rh(D)-отрицательных лиц среди пациентов составило 94,39 и 5,6% соответственно. Для доноров это соотношение составило 92,43% Rh(D)-положительных и 7,57% Rh(D)-отрицательных (табл. 2).
Полученные нами данные выявили значительно больший процент антигена D вариантного у пациентов. Возможно, это является результатом ослабления антигена D при заболеваниях. Изучение антигенного состава эритроцитов кадровых доноров и пациентов показало, что наиболее распространенным фенотипом среди Rh(D)-положительных лиц является фенотип CCDee-, далее следуют фенотипы CcDee-, CcDEe- и ccDEe- (табл. 3).
3).
Наибольшую опасность представляют в трансфузионном отношении гомозиготные сочетания антигенов системы Резус (СС, сc, ЕЕ и ее), которые встречаются у 86% резус-положительных (D+). Всем донорам и реципиентам независимо от групповой и резус- принадлежности предварительно проводился скрининг на наличие антиэритроцитарных антител. При оценке антиэритроцитарных антител у 13,3% пациентов обнаружены иммунные антитела различной специфичности. В связи с тем, что среди контингента обследованных в лаборатории лиц превалировали беременные, входившие в группу риска по аллоиммунизации, а также больные получившие многократные трансфузии, – индекс сенсибилизации у пациентов оказался высоким. Антитела к антигенам эритроцито системы Резус появляются в организме в результате трансфузий эритроцитов доноров, содержащих антигены, отсутствующие у реципиента, а также при иммунизации матери эритроцитами плода. Частота встречаемости аллоантител к антигенам эритроцитов системы Резус различна и определяется иммуногенностью антигена и частотой встречаемости в популяции. В 31% случаев антитела не идентифицировали, что связано с сочетанной специфичностью антител или наличием антител к широко распространенным в популяции антигенам. При изучении частоты аллосенсибилизации среди RH(D)-положительных и RH(D)-отрицательных лиц, было обнаружено, что RH(D)-отрицательные люди иммунизируются в 2–2,5 раза чаще, чем RH(D)-положительные. Антитела к минорным антигенам эритроцитов чаще встречаются у резус-положительных лиц, чем у резус-отрицательных. У доноров встречаемость аллоантител составила 0,46% (табл. 4).
В 31% случаев антитела не идентифицировали, что связано с сочетанной специфичностью антител или наличием антител к широко распространенным в популяции антигенам. При изучении частоты аллосенсибилизации среди RH(D)-положительных и RH(D)-отрицательных лиц, было обнаружено, что RH(D)-отрицательные люди иммунизируются в 2–2,5 раза чаще, чем RH(D)-положительные. Антитела к минорным антигенам эритроцитов чаще встречаются у резус-положительных лиц, чем у резус-отрицательных. У доноров встречаемость аллоантител составила 0,46% (табл. 4).
Проблема гемолитической болезни новорожденных (ГБН) для города Астана актуальна. Наиболее часто ГБН развивается вследствие конфликта по антигенам системы Резус и АВ0 (табл. 5).
методов
метод
НАЗАД
Антропологические типы, представленные на этом сайте, получены в результате качественного анализа множества различных антропологических исследований, в основном написанных в 20-м веке, в основном на немецком, английском, французском, итальянском, русском, шведском, венгерском или польском языках. Классические антропологические типологии в одних случаях были похожи друг на друга, а в других сильно отличались друг от друга. Для получения типов, изображенных на этом сайте, сравнивались типовые описания разных авторов, и в случае сходства несколько типов объединялись в один, а в случае различия сохранялись исходные типы. Чтобы понять, почему был выбран именно этот подход, какие существуют альтернативы, как классические исследования 20-го века определили их типы и как они соотносятся с современными, часто филогенетическими исследованиями 21-го века, здесь дается методологический обзор. Классические антропологические типологии в одних случаях были похожи друг на друга, а в других сильно отличались друг от друга. Для получения типов, изображенных на этом сайте, сравнивались типовые описания разных авторов, и в случае сходства несколько типов объединялись в один, а в случае различия сохранялись исходные типы. Чтобы понять, почему был выбран именно этот подход, какие существуют альтернативы, как классические исследования 20-го века определили их типы и как они соотносятся с современными, часто филогенетическими исследованиями 21-го века, здесь дается методологический обзор. В большинстве случаев на этом сайте приоритет отдается интегральному подходу. Поскольку колонизация и глобализация быстро изменили антропологический состав многих регионов мира, все карты и описания относятся к 1500 году , до того, как эти процессы начались в больших масштабах. Иллюстрировать нынешнее современное состояние разнообразия было бы противоречиво, поскольку в настоящее время любой тип может появиться в любой точке мира. Далее рассматривается, откуда берутся определения антропологических типов, являются ли они четкими биологическими подразделениями или же они «падают с неба» и являются совершенно случайными, рукотворными, даже политическими определениями. Вся информация является результатом частного хобби-проекта и не основана на финансировании каких-либо исследований. Хотя сайт старается быть максимально точным, некоторые результаты носят спекулятивный характер и могут содержать ошибки. Если вы найдете ошибки, пожалуйста, свяжитесь со мной. |
 Кроме того, многие смешанные группы еще не стабилизированы, т.е. члены одной семьи могут сильно различаться. Стабилизация обычно занимает несколько столетий.
Кроме того, многие смешанные группы еще не стабилизированы, т.е. члены одной семьи могут сильно различаться. Стабилизация обычно занимает несколько столетий.1.) Численный подход
Численный подход анализирует комбинации антропометрических признаков на индивидуальном уровне (в отличие от популяционного подхода). Эта методология также является основой интегрального подхода и популяционного подхода и была частью анализа для большинства антропологов. Однако, в отличие от, например, интегральный подход, численный подход только использует метрики для определения типа. Это имеет то преимущество, что никакие «человеческие предубеждения» не изменяют результаты, например. субъективное впечатление, которое некоторые фенотипы производят на человека-наблюдателя. Кроме того, анализ данных может выявить взаимосвязи, которые иначе невозможно наблюдать. Таким образом, числовые типы являются исключительно статистическим результатом. Базовая схема классификации европейских типов основана на числовом подходе: Эта методология также является основой интегрального подхода и популяционного подхода и была частью анализа для большинства антропологов. Однако, в отличие от, например, интегральный подход, численный подход только использует метрики для определения типа. Это имеет то преимущество, что никакие «человеческие предубеждения» не изменяют результаты, например. субъективное впечатление, которое некоторые фенотипы производят на человека-наблюдателя. Кроме того, анализ данных может выявить взаимосвязи, которые иначе невозможно наблюдать. Таким образом, числовые типы являются исключительно статистическим результатом. Базовая схема классификации европейских типов основана на числовом подходе:Таблица 1: Пример для европейских типов на основе числового подхода по сочетанию трех антропометрических признаков (Лебцельтер, 1929):
Головной указатель не менее 81 считается короткоголовым, поэтому любой высокий светловолосый человек с головным указателем 80,9 будет классифицирован как нордид с использованием этого подхода, основанного на трех признаках. Рисунок 1: Количество возможных комбинаций типов дает определенное количество антропометрических признаков, которые имеют два или три проявления признака: |

 С индексом 81 он был бы Норидом, если не учитывать другие антропометрические особенности.
С индексом 81 он был бы Норидом, если не учитывать другие антропометрические особенности. Однако рассмотрение только 5, 10 или даже 50 антропометрических признаков будет недостаточным для описания такого сложного организма, как человек, они могут отразить лишь небольшую часть фенотипа. Кроме того, погрешность измерения или разные способы измерения (например, существует несколько методов получения носового индекса) или метрические признаки, измененные окружающей средой, легко могут повлиять на классификацию. Человеческая предвзятость также легко вносится при выборе данных, полученных для анализа.
Однако рассмотрение только 5, 10 или даже 50 антропометрических признаков будет недостаточным для описания такого сложного организма, как человек, они могут отразить лишь небольшую часть фенотипа. Кроме того, погрешность измерения или разные способы измерения (например, существует несколько методов получения носового индекса) или метрические признаки, измененные окружающей средой, легко могут повлиять на классификацию. Человеческая предвзятость также легко вносится при выборе данных, полученных для анализа.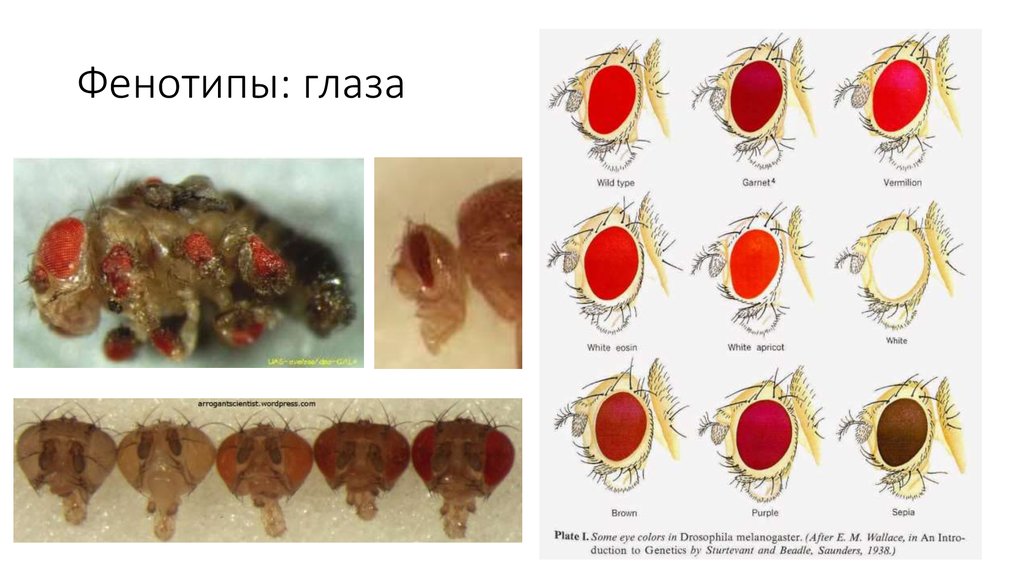
2.) Интегральный подход
| Интегральный подход является предпочтительным для типов, представленных на этом сайте. Только в том случае, если у антропологов, использующих интегральный подход, было мало информации о конкретном месте в мире, для региона включались типы других подходов. Такие подходы были распространены в немецкой, итальянской и французской антропологических школах. Интегральный подход родственен численному подходу, точнее, строится на нем. Он также использует статистические комбинации антропометрических признаков (см. таблицу 1), но не полностью полагается на них. Вместо этого он добавляет большое количество дополнительной качественной информации. Это включает в себя качественную оценку физических признаков и того, как они выглядят для глаза человека-наблюдателя, особенно особенностей мягких тканей человека, которые трудно измерить. Рисунок 2: Динарид или Нордид? (сравните с таблицей 1) Для любого определенного набора идеальных типов (типология) люди будут группироваться в три категории: 2.) «Смешанные» особи, которые соответствуют (линейной) комбинации двух или более типов. 3.) Лица, которые не могут быть охвачены типологией. Чтобы найти набор типов, интегральный антрополог, например, может попытаться максимизировать количество людей в группе 1 и минимизировать количество людей в группе 3. В идеале определенный тип должен доминировать в определенных популяциях, социальных группах или семьях. и быть характерным для них. Интегральный подход сталкивается с рядом проблем. Большой объем информации, которую необходимо рассмотреть, может быть слишком сложным, чтобы найти набор возможных типов, который лучше всего описывает человеческое разнообразие. В целом интегральный подход является очень мощным инструментом для демонстрации разнообразия. Поэтому он был выбран для этого сайта. В отличие от некоторых классических антропологов, которые продвигали свои выводы как определенные по разным причинам, здесь они рассматриваются просто как фенотипическая модель. |
 Интегральный подход также учитывает исторические факты, т.е. о старых миграциях, семейной истории человека и окружающей среде, в которой живет человек, потому что они могут изменить фенотипы. Это также имеет значение, если тип сохраняется в семьях, социальных группах или популяциях, например. семейный или деревенский. Только после учета всей возможной информации производится классификация.
Интегральный подход также учитывает исторические факты, т.е. о старых миграциях, семейной истории человека и окружающей среде, в которой живет человек, потому что они могут изменить фенотипы. Это также имеет значение, если тип сохраняется в семьях, социальных группах или популяциях, например. семейный или деревенский. Только после учета всей возможной информации производится классификация. Такой человек будет классифицирован как динарид при строгом числовом подходе, основанном на таблице 1, в которой используются три черты. На рисунке ясно видны значительные различия между Nordoid и Dinarid, хотя они были бы одинаковыми в приведенном выше примере численного подхода. Это связано с тем, что трех признаков обычно слишком мало, чтобы зафиксировать человеческий фенотип.
Такой человек будет классифицирован как динарид при строгом числовом подходе, основанном на таблице 1, в которой используются три черты. На рисунке ясно видны значительные различия между Nordoid и Dinarid, хотя они были бы одинаковыми в приведенном выше примере численного подхода. Это связано с тем, что трех признаков обычно слишком мало, чтобы зафиксировать человеческий фенотип.
 В целом число теоретически возможных определений типов, которые могут быть включены в типологию, равно 9.0008 почти бесконечное , хотя лишь немногие из потенциальных типологий действительно хорошо отражают человеческое разнообразие. Могут существовать аналогичные типологии аналогичного значения, которые также описывают разнообразие. Таким образом, определения типов интегральных подходов иногда различаются, особенно в деталях. Таким образом, любая типология больше похожа на модель, которая существует в большинстве наук в качестве полезного инструмента — как упрощение реальности, которое никогда не может охватить каждую деталь реального мира. Кроме того, впечатление, которое человек может произвести на человека-наблюдателя для качественной оценки, может быть предвзятым, поскольку такая оценка зависит от навыков, личности, мотивации и опыта наблюдателя. Другая проблема заключается в том, что исторические данные и антропометрические измерения могут содержать ошибки. В настоящее время можно было бы частично заменить информацию из исторических документов и сказок (например, о старых миграциях) генетической информацией и иметь больше и более качественные антропометрические данные.
В целом число теоретически возможных определений типов, которые могут быть включены в типологию, равно 9.0008 почти бесконечное , хотя лишь немногие из потенциальных типологий действительно хорошо отражают человеческое разнообразие. Могут существовать аналогичные типологии аналогичного значения, которые также описывают разнообразие. Таким образом, определения типов интегральных подходов иногда различаются, особенно в деталях. Таким образом, любая типология больше похожа на модель, которая существует в большинстве наук в качестве полезного инструмента — как упрощение реальности, которое никогда не может охватить каждую деталь реального мира. Кроме того, впечатление, которое человек может произвести на человека-наблюдателя для качественной оценки, может быть предвзятым, поскольку такая оценка зависит от навыков, личности, мотивации и опыта наблюдателя. Другая проблема заключается в том, что исторические данные и антропометрические измерения могут содержать ошибки. В настоящее время можно было бы частично заменить информацию из исторических документов и сказок (например, о старых миграциях) генетической информацией и иметь больше и более качественные антропометрические данные. Такая генетически расширенная интегральная типология может быть разработана путем максимизации группы 1 и минимизации группы 3 после сбора данных. В качестве альтернативы количество переменных может быть уменьшено с помощью анализа основных компонентов и аналогичных методов. Поскольку у классических антропологов такой информации не было, вполне возможно, что некоторые из их типов устарели бы при современном подходе.
Такая генетически расширенная интегральная типология может быть разработана путем максимизации группы 1 и минимизации группы 3 после сбора данных. В качестве альтернативы количество переменных может быть уменьшено с помощью анализа основных компонентов и аналогичных методов. Поскольку у классических антропологов такой информации не было, вполне возможно, что некоторые из их типов устарели бы при современном подходе.3.) Население
Проблемы неоднозначности других типологических подходов можно преодолеть с помощью популяционного подхода. Этот подход был особенно распространен во второй половине 20-го века и обычно использовался русской антропологической школой, а также некоторыми французскими, немецкими и другими исследователями. Вместо того, чтобы рассматривать отдельных лиц, популяционный подход рассматривает средние показатели комплексов населения или языковых групп. Эти группы могут содержать широкий спектр фенотипов. Явным преимуществом популяционного подхода является то, что популяции или языковые группы часто могут быть четко определены, следовательно, существует уникальное, четкое решение. Кроме того, его применимость очень высока. Может быть включено множество различных метрик, и для результирующей системы типов не существует ограничений, как в двух подходах, показанных выше. Теоретически каждый популяционный комплекс хорошо узнаваем даже при оценке 1000 антропометрических признаков. Первые компьютеры 20-го века могли обрабатывать усредненные показатели населения гораздо проще, чем отдельные наборы данных. Таким образом, он добавил много новых выводов в исследование. Образовавшиеся комплексы населения, т.е. Балтийский, Западно-Балтийский и т. д. обычно не идентичны типам других подходов, хотя могут быть и схожими. Вместо того, чтобы рассматривать отдельных лиц, популяционный подход рассматривает средние показатели комплексов населения или языковых групп. Эти группы могут содержать широкий спектр фенотипов. Явным преимуществом популяционного подхода является то, что популяции или языковые группы часто могут быть четко определены, следовательно, существует уникальное, четкое решение. Кроме того, его применимость очень высока. Может быть включено множество различных метрик, и для результирующей системы типов не существует ограничений, как в двух подходах, показанных выше. Теоретически каждый популяционный комплекс хорошо узнаваем даже при оценке 1000 антропометрических признаков. Первые компьютеры 20-го века могли обрабатывать усредненные показатели населения гораздо проще, чем отдельные наборы данных. Таким образом, он добавил много новых выводов в исследование. Образовавшиеся комплексы населения, т.е. Балтийский, Западно-Балтийский и т. д. обычно не идентичны типам других подходов, хотя могут быть и схожими. Обычно внутри каждого популяционного комплекса выделяют несколько типов. На рис. 3 приведен пример. В верхнем ряду показаны локальные разновидности, выделенные с помощью интегрального подхода, во втором ряду приведены примеры популяционных комплексов, которые эти типы могли образовать. Обычно внутри каждого популяционного комплекса выделяют несколько типов. На рис. 3 приведен пример. В верхнем ряду показаны локальные разновидности, выделенные с помощью интегрального подхода, во втором ряду приведены примеры популяционных комплексов, которые эти типы могли образовать.Рисунок 3: Соотношение популяционных комплексов и антропологических типов интегрального подхода: Рисунок 3 также показывает один недостаток популяционного подхода: все три изображенных комплекса выглядят одинаково. Это связано с тем, что по мере агрегирования разнообразия фенотипов внутри популяции может быть потеряно много информации. Популяции могут быть разнообразными, социальные группы или группы предков внутри популяции могут существенно различаться, и это не всегда известно исследователю. Определенная социальная группа может быть больше похожа на жителей другого региона, чем на сожительствующие социальные группы. Если взять только среднее значение и дисперсию нескольких показателей по популяциям, возможно, будет отброшено большое количество информации, доступной только на индивидуальном уровне. Рис. 4: Плотность распределения вероятностей оценочного роста мужчин для места проживания нордидов и лаппидов в Северной Скандинавии: |
 В качестве примера на рисунке 4 показана оценочная функция плотности вероятности одного антропометрического признака, мужского роста, для места в Северной Скандинавии, населенного нордскими и саамскими людьми. Нордиды в среднем значительно выше саамов. Таким образом, функция имеет два пика. Подобные явления обнаруживаются во многих местах мира и во многих различных антропометрических характеристиках. Эта информация не может быть получена комплексом населения, если различные группы внутри населения не разделены. Это особенно невыгодно в очень разных странах, таких как Индия, где кастовая система порождала большое разнообразие образов.
В качестве примера на рисунке 4 показана оценочная функция плотности вероятности одного антропометрического признака, мужского роста, для места в Северной Скандинавии, населенного нордскими и саамскими людьми. Нордиды в среднем значительно выше саамов. Таким образом, функция имеет два пика. Подобные явления обнаруживаются во многих местах мира и во многих различных антропометрических характеристиках. Эта информация не может быть получена комплексом населения, если различные группы внутри населения не разделены. Это особенно невыгодно в очень разных странах, таких как Индия, где кастовая система порождала большое разнообразие образов. Однако популяционный подход остается наиболее надежным в практическом смысле.
Однако популяционный подход остается наиболее надежным в практическом смысле.4.) Раса как социальный конструкт
Другой подход, очень распространенный сегодня, состоит в том, чтобы определить антропологические группы или «расы» как социальную конструкцию. Это имеет мало общего с тремя подходами, проиллюстрированными выше, и еще меньше с филогенетическими отношениями. Критерии классификации различаются от страны к стране и меняются со временем. Человек, считающийся «черным» в Соединенных Штатах, может не рассматриваться как черный в других частях мира, человек может считаться «белым» в Уганде или даже в Австралии. В подходе социальных конструктов классификация людей часто связана с историей страны. Типичными примерами являются «правила одной капли». Например. если американец имеет 1/8 африканского происхождения, он будет классифицирован как афроамериканец, даже если фенотипически и генетически он может быть в основном европейцем. И наоборот, человек с 1/8 европейского происхождения не будет считаться в этой системе «белым» или «европейцем». Большую роль играет и то, как человек себя ведет, где он вырос и как себя идентифицирует. У некоторых людей может быть происхождение, при котором они могут выбирать свою «расу», а у людей с другим происхождением — нет. Этот подход также применялся к коренным американцам в США, коренным австралийцам или даже айнам в Японии. Другой характеристикой подхода социальных конструктов является чрезмерный акцент на единственной антропометрической характеристике, часто на цвете кожи. Например. Суданиды. Эфиопидов, индо-меланидов, негритидов и австралидов можно охарактеризовать просто как «черных», хотя они сильно различаются генетически, исторически и даже антропометрически. У них просто есть хотя бы одна общая антропометрическая черта. Эквивалентным было бы объединение всех широколицых людей в одну «расу» (чего никогда не делалось). В некоторых случаях политические идеологии играют большую роль в подходе социальных конструктов, несколько десятилетий назад аналогичные методы в антисемитских странах Европы использовались для идентификации евреев. И наоборот, человек с 1/8 европейского происхождения не будет считаться в этой системе «белым» или «европейцем». Большую роль играет и то, как человек себя ведет, где он вырос и как себя идентифицирует. У некоторых людей может быть происхождение, при котором они могут выбирать свою «расу», а у людей с другим происхождением — нет. Этот подход также применялся к коренным американцам в США, коренным австралийцам или даже айнам в Японии. Другой характеристикой подхода социальных конструктов является чрезмерный акцент на единственной антропометрической характеристике, часто на цвете кожи. Например. Суданиды. Эфиопидов, индо-меланидов, негритидов и австралидов можно охарактеризовать просто как «черных», хотя они сильно различаются генетически, исторически и даже антропометрически. У них просто есть хотя бы одна общая антропометрическая черта. Эквивалентным было бы объединение всех широколицых людей в одну «расу» (чего никогда не делалось). В некоторых случаях политические идеологии играют большую роль в подходе социальных конструктов, несколько десятилетий назад аналогичные методы в антисемитских странах Европы использовались для идентификации евреев. Подход социальных конструктов не используется на этом сайте, потому что он не может описать биологическое разнообразие человека научным способом. Подход социальных конструктов не используется на этом сайте, потому что он не может описать биологическое разнообразие человека научным способом. |
5.) Филогенетический подход
Очень мощным научным подходом к изучению человеческого разнообразия является филогенетический подход. Вместо рассмотрения фенотипических и антропометрических признаков он изучает генетический состав людей, часто на основе данных молекулярного секвенирования. Термин «филогенетика» происходит от греческих терминов «фил» и «филон», обозначающих «племя», «клан», «расу». С развитием генетических методов в конце 20-го и начале 21-го века этот подход стал распространенным методом исследования биологического разнообразия человека, хотя 100 лет назад уже существовали филогенетические деревья, которые имели сходство с современными деревьями. Как правило, разнообразие генетических признаков оценивается, а затем с помощью статистического анализа объединяется в основные компоненты или аналогичные факторы. Филогенетический подход лучше всего подходит для изучения древних миграций, времени расхождения человеческих популяций и степени их смешения. Однако он сталкивается с теми же статистическими проблемами, что и численный подход, когда речь идет о сопоставимости и качестве данных. В большинстве случаев собранная генетическая информация будет описывать только часть всего биологического организма, данные могут содержать ошибки, а методы измерения не всегда сопоставимы. Это может привести к необъективным результатам, поскольку сложный организм не рассматривается полностью, а многие ненаблюдаемые характеристики и гены игнорируются. Кроме того, применяемые статистические методы имеют ограничения и особые требования к данным, которые не всегда могут быть удовлетворены. Другая слабость заключается в том, что филогенетический подход не учитывает фенотипические различия (см. ниже). Однако, несмотря на эти недостатки, возможность использования филогенетического подхода для изучения древних миграций человека остается большой. Филогенетический подход лучше всего подходит для изучения древних миграций, времени расхождения человеческих популяций и степени их смешения. Однако он сталкивается с теми же статистическими проблемами, что и численный подход, когда речь идет о сопоставимости и качестве данных. В большинстве случаев собранная генетическая информация будет описывать только часть всего биологического организма, данные могут содержать ошибки, а методы измерения не всегда сопоставимы. Это может привести к необъективным результатам, поскольку сложный организм не рассматривается полностью, а многие ненаблюдаемые характеристики и гены игнорируются. Кроме того, применяемые статистические методы имеют ограничения и особые требования к данным, которые не всегда могут быть удовлетворены. Другая слабость заключается в том, что филогенетический подход не учитывает фенотипические различия (см. ниже). Однако, несмотря на эти недостатки, возможность использования филогенетического подхода для изучения древних миграций человека остается большой. На рис. 5 показано сильно упрощенное филогенетическое древо современного человека. На рис. 5 показано сильно упрощенное филогенетическое древо современного человека. |
Рисунок 5: Упрощенное филогенетическое дерево человека для выбранных антропологических типов согласно текущим исследованиям:
Филогенетическое древо показывает общего предка Homo sapiens и неандертальцев, живших около полумиллиона лет назад в Африке. Неандертальцы, покинувшие Африку за несколько сотен тысяч лет до Homo sapiens, приспособились к прохладному климату ледникового периода Европы и Азии, развив коренастое тело, очень большие носы и более светлую кожу. Homo sapiens начал расходиться на современные разновидности около 150 000 лет до н. э. в Африке. Ранние Homo sapiens, вероятно, жили в сухом тропическом климате Африки (во время ледникового периода тропики были менее влажными, чем сегодня). Таким образом, носы, вероятно, были лишь умеренно широкими (см. взаимосвязь между шириной носа и влажностью), а кожа, вероятно, была (темно) коричневой. Койсан отделился первым от всех остальных людей. Общий предок всех некойсанских групп развил разные разновидности: протосуданиды, протонилотиды и протоэтиопиды. Последняя группа начала мигрировать из Африки около 60 000 лет назад и смешалась с неандертальцами и, возможно, другими людьми, такими как денисовцы. Когда люди мигрировали на север, их кожа стала светлее, часто достигая светлого или среднего коричневого тона. Фенотипы быстро начали меняться в новых условиях из-за адаптивного давления, естественного отбора и культурных изменений, которые привнесли новые условия. Северо-восточные азиаты приспособились к континентальному климату с ледяными зимами, европейцы к земледельческой жизни в облачном умеренном климате. Обе адаптации привели от скромной к сильной депигментации, более узким носам и более коротким конечностям. Некоторые группы мигрировали через Южную Азию в Австралию и Меланезию (например, предки букаидов). Хотя букаиды генетически ближе к европейцам и выходцам из Восточной Азии, они выглядят относительно похожими на бантуидов и многих других африканцев к югу от Сахары из-за их адаптации к аналогичному влажному и тропическому климату. Койсан отделился первым от всех остальных людей. Общий предок всех некойсанских групп развил разные разновидности: протосуданиды, протонилотиды и протоэтиопиды. Последняя группа начала мигрировать из Африки около 60 000 лет назад и смешалась с неандертальцами и, возможно, другими людьми, такими как денисовцы. Когда люди мигрировали на север, их кожа стала светлее, часто достигая светлого или среднего коричневого тона. Фенотипы быстро начали меняться в новых условиях из-за адаптивного давления, естественного отбора и культурных изменений, которые привнесли новые условия. Северо-восточные азиаты приспособились к континентальному климату с ледяными зимами, европейцы к земледельческой жизни в облачном умеренном климате. Обе адаптации привели от скромной к сильной депигментации, более узким носам и более коротким конечностям. Некоторые группы мигрировали через Южную Азию в Австралию и Меланезию (например, предки букаидов). Хотя букаиды генетически ближе к европейцам и выходцам из Восточной Азии, они выглядят относительно похожими на бантуидов и многих других африканцев к югу от Сахары из-за их адаптации к аналогичному влажному и тропическому климату. Рисунок 5 показывает, что при относительно постоянных условиях в доисторической Африке фенотипы менялись с умеренной скоростью, в то время как они быстро менялись в различных новых условиях за пределами Африки. Рисунок 5 показывает, что при относительно постоянных условиях в доисторической Африке фенотипы менялись с умеренной скоростью, в то время как они быстро менялись в различных новых условиях за пределами Африки.По крайней мере два разных процесса определяют человеческое разнообразие: генетическая дивергенция и фенотипическая дивергенция. Основной движущей силой генетической дивергенции являются стохастические генетические мутации, которые происходят с течением времени, в то время как основной движущей силой фенотипической дивергенции является адаптивное давление в новых условиях, культуре и возникающих в результате процессах естественного отбора. Разделенные популяции при сходных обстоятельствах долгое время будут оставаться похожими из-за одинакового адаптивного давления и, возможно, даже конвергентной эволюции. Для различий между людьми важны оба процесса. Это показано ниже. |
Почему важны фенотипы?
В предыдущем разделе было показано, что филогенетический подход очень полезен для изучения человеческих миграций и генетической дивергенции. Однако различия у людей обусловлены не только генетической дивергенцией, но и фенотипической дивергенцией, вызванной адаптивным давлением. В ходе эволюции новый вид может быть создан любым из этих процессов, в большинстве случаев комбинацией обоих. Если не будет фенотипической дивергенции, а будет длительный процесс генетической дивергенции, в разных популяциях будет развиваться слишком много генетических различий, чтобы продолжать производить жизнеспособное потомство и развиваться в новые виды. Если не будет генетической дивергенции, а будет сильный естественный отбор и адаптивное давление на одну или обе популяции, их анатомия станет настолько другой, что они также перестанут скрещиваться. До видообразования обычно развиваются разные подвиды или «расы». Не существует единого определения, с какого момента разные фенотипы и/или генотипы могут рассматриваться как разные подвиды. Это иллюстрируется «серой зоной» на рис. 6, которая изображает два процесса (гораздо более подробное описание фенотипической и генетической дивергенции в эволюции см. Однако различия у людей обусловлены не только генетической дивергенцией, но и фенотипической дивергенцией, вызванной адаптивным давлением. В ходе эволюции новый вид может быть создан любым из этих процессов, в большинстве случаев комбинацией обоих. Если не будет фенотипической дивергенции, а будет длительный процесс генетической дивергенции, в разных популяциях будет развиваться слишком много генетических различий, чтобы продолжать производить жизнеспособное потомство и развиваться в новые виды. Если не будет генетической дивергенции, а будет сильный естественный отбор и адаптивное давление на одну или обе популяции, их анатомия станет настолько другой, что они также перестанут скрещиваться. До видообразования обычно развиваются разные подвиды или «расы». Не существует единого определения, с какого момента разные фенотипы и/или генотипы могут рассматриваться как разные подвиды. Это иллюстрируется «серой зоной» на рис. 6, которая изображает два процесса (гораздо более подробное описание фенотипической и генетической дивергенции в эволюции см. , например, Winker, 2009).). , например, Winker, 2009).).Рисунок 6: Фенотипическая и генетическая дивергенция: Чтобы проиллюстрировать оба процесса, можно нарисовать несколько пар фенотипов с рисунка 5 на рисунок 6: Рисунок 7: Фенотипическая и генетическая дивергенция для выбранных пар типов из рисунка 5: Рисунок 7 иллюстрирует фенотипическую и генетическую дивергенцию для трех типов с рисунка 5: нордид, букаид и неандертальцев. Между неандертальцами и нордидами генетические, а также фенотипические расхождения относительно велики, некоторые исследователи считают неандертальцев отдельным видом. Между нордидами и букаидами фенотипические различия меньше, чем у неандертальцев, но различия между нордидами и букаидами легко распознаются по многим антропометрическим признакам, оба показывают совершенно разную адаптацию. Однако эти два типа, вероятно, разделились только 40 000–60 000 лет назад, что является относительно поздним периодом эволюции. Раскол между бантуидами и букаидами произошел раньше (см. Таким образом, фенотипические, а также генетические различия объясняют разнообразие людей. Человеческое разнообразие может иметь значение для здоровья, питания, общества, спорта и навыков. Текущее исследование добавляет много информации о генетических различиях, но мало о фенотипических различиях. Однако на протяжении большей части палеолита, когда основные современные ветви Homo sapiens расходились, люди были тесно связаны с климатом и природой, в которых они жили, их фенотипы были зеркалом окружающей их среды. Таким образом, Humanphenotypes пытается закрыть этот пробел, предоставляя типологическую модель, которая обобщает доступную информацию о фенотипических различиях между людьми. Данные и типы в основном изучались в течение 20 века. |
 рис. 5), но все же оба типа похожи друг на друга. Это связано с тем, что оба живут в относительно похожем климате, и адаптивные нагрузки были одинаковыми. Таким образом, хотя генетические различия между букаидами и бантуидами больше, фенотипическая разница невелика.
рис. 5), но все же оба типа похожи друг на друга. Это связано с тем, что оба живут в относительно похожем климате, и адаптивные нагрузки были одинаковыми. Таким образом, хотя генетические различия между букаидами и бантуидами больше, фенотипическая разница невелика.