Dux architeuthis: ADW: Architeuthis dux: INFORMATION
Architeuthis dux Steenstrup, 1857
Architeuthis dux Steenstrup, 1857
К сожалению, GBIF не работает без разрешения на запуск JavaScript.
Наш сайт сайт обнаружил, что Вы используете устаревший и небезопасный браузер. Это не позволит Вам использовать данный сайт. Пожалуйста, установите современный браузер.
- Dataset
- GBIF Backbone Taxonomy
- Rank
- SPECIES
- Published in
- Steenstrup J.
 (1857). Oplysninger om Atlanterhavets colossale Blaeksprutter. <em>Forhandlinger ved de Skandinaviske Naturforskeres.</em> 7 (1856): 182-185.
(1857). Oplysninger om Atlanterhavets colossale Blaeksprutter. <em>Forhandlinger ved de Skandinaviske Naturforskeres.</em> 7 (1856): 182-185.
 (1857). Oplysninger om Atlanterhavets colossale Blaeksprutter. <em>Forhandlinger ved de Skandinaviske Naturforskeres.</em> 7 (1856): 182-185.
(1857). Oplysninger om Atlanterhavets colossale Blaeksprutter. <em>Forhandlinger ved de Skandinaviske Naturforskeres.</em> 7 (1856): 182-185.http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons. org/licenses/by-nc/4.0/
org/licenses/by-nc/4.0/
http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons.org/licenses/by/4.0/
http://creativecommons. org/licenses/by-nc/4.0/
org/licenses/by-nc/4.0/
http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons. org/licenses/by-nc/4.0/
org/licenses/by-nc/4.0/
http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons. org/licenses/by-nc/4.0/
org/licenses/by-nc/4.0/
http://creativecommons.org/publicdomain/zero/1.0/
Usage Conditions Apply
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons. org/publicdomain/zero/1.0/
org/publicdomain/zero/1.0/
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons. org/publicdomain/zero/1.0/
org/publicdomain/zero/1.0/
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons. org/publicdomain/zero/1.0/
org/publicdomain/zero/1.0/
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons. org/publicdomain/zero/1.0/
org/publicdomain/zero/1.0/
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons.org/licenses/by/4.0/
http://creativecommons.org/licenses/by/4.0/
http://creativecommons. org/licenses/by/4.0/
org/licenses/by/4.0/
http://creativecommons.org/licenses/by/4.0/
http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons. org/licenses/by-nc/4.0/
org/licenses/by-nc/4.0/
http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons.org/licenses/by-nc/4.0/
http://creativecommons. org/publicdomain/zero/1.0/
org/publicdomain/zero/1.0/
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons. org/publicdomain/zero/1.0/
org/publicdomain/zero/1.0/
http://creativecommons.org/publicdomain/zero/1.0/
http://creativecommons. org/publicdomain/zero/1.0/
org/publicdomain/zero/1.0/
http://creativecommons.org/licenses/by/4.0/
http://creativecommons.org/licenses/by/4.0/
http://creativecommons.org/licenses/by/4.0/
Classification
- kingdom
- Animalia
- phylum
- Mollusca
- class
- Cephalopoda
- order
- Oegopsida
- family
- Architeuthidae
- genus
- Architeuthis
- species
- Architeuthis dux
Name
- Synonyms
- Architeuthis clarkei Robson, 1933
- Architeuthis hartingii (Verrill, 1875)
- Architeuthis harveyi (Kent, 1874)
- Architeuthis japonica Pfeffer, 1912
- Architeuthis kirkii Robson, 1887
- Architeuthis longimanus Kirk, 1888
- Architeuthis martensii (Hilgendorf, 1880)
- Architeuthis monachus Steenstrup
- Architeuthis nawaji Cadenat, 1935
- Architeuthis physeteris (Joubin, 1900)
- Architeuthis princeps Verrill, 1875
- Architeuthis sanctipauli (Vélain, 1877)
- Architeuthis stockii (Kirk, 1882)
- Architeuthis verrilli Kirk, 1882
- Dinoteuthis proboscideus More, 1875
- Dubioteuthis physeteris Joubin, 1900
- Loligo hartingii Verrill, 1875
- Megaloteuthis harveyi Kent, 1874
- Megateuthus martensii Hilgendorf, 1880
- Mouchezis sanctipauli Vélain, 1877
- Homonyms
- Architeuthis dux Steenstrup, 1857
- Common names
- Atlantic giant squid in Английский
- Atlantischer Riesenkalmar in Немецкий
- Dà-Wàng-Yóu in Китайский
- Kæmpeblæksprutte in Датский
- calmar géant in Французский
- giant squid in Английский
- jättebläckfiskar in Шведский
- Гигантский кальмар in Русский
- jättebläckfisk in Шведский
- Atlantic giant squid in Английский
- Atlantischer Riesenkalmar in Немецкий
- Dà-Wàng-Yóu in Китайский
- calmar géant in Французский
- giant oceanic squid in Английский
- giant squid in Английский
- jättebläckfiskar in Шведский
- Гигантский кальмар in Русский
Bibliographic References
- Harting, P. , 1860: Description de quelques fragments de deux Cephalopodes gigantesques. Verhandelingen der Koninklijke Akademie van Wetenschappen, Amsterdam, 9 (1). 16.
- Ibáñez et al. 2009
- Judkins, H. L., M. Vecchione, and C. F. E. Roper. 2009. Cephalopoda (Mollusca) of the Gulf of Mexico, Pp. 701–709 in Felder, D.L. and D.K. Camp (eds.), Gulf of Mexico–Origins, Waters, and Biota. Biodiversity. Texas A&M Press, College Station, Texas.
- Karlsson, Anna (2015-03-23 00:00:00) Mollusker
- Kristensen, T. K. and J. Knudsen, 1983: A catalogue of the type specimens of Cephalopoda (Mollusca) in the Zoological Museum, University of Copenhagen. Steenstrupia, 9 (10). 217-227.
- Lu, C.C. & Chung, W.S. (2017). <em>Guide to the cephalopods of Taiwan</em>. National Museum of Natural Science, Taichung, Taiwan, 560 pp. ISBN 978-986-05-2569-4.
- Malmberg & Lundin (2019-02-01 16:02:00) Svenska bläckfisknamn, antagna 190123 av Kommittén för svenska djurnamn
- Marshall, B. A. (1996). Molluscan name-bearing types in the Museum of New Zealand Te Papa Tongarewa. <em>Tuhinga: Records of the Museum of New Zealand Te Papa Tongarewa.</em> 9: 1-85.
- Nesis, K. N., 1987: null. Cephalopods of the World: Squids, Cuttlefishes, Octopuses, and Allies. 351.
- Roper, C.F.E. & Jereb, P. (2010). Family Architeuthidae. <i>In</i> P. Jereb & C.F.E. Roper, eds. <i>Cephalopods of the world. An annotated and illustrated catalogue of species known to date. Volume 2. Myopsid and Oegopsid Squids</i>. FAO Species Catalogue for Fishery Purposes. No. 4, Vol. 2. Rome, FAO. <em>pp. 121-123.</em>
- Spencer, H.G., Marshall, B.A. & Willan, R.C. (2009). Checklist of New Zealand living Mollusca. Pp 196-219. <em>in: Gordon, D.P. (ed.) New Zealand inventory of biodiversity. Volume one. Kingdom Animalia: Radiata, Lophotrochozoa, Deuterostomia.</em> Canterbury University Press, Christchurch.
- Steenstrup, J. (1857). Oplysninger om Atlanterhavets colossale Blaeksprutter. <em>Forhandlinger ved de Skandinaviske Naturforskeres.</em> 7 (1856): 182-185.
- Steenstrup, J. , 1857: Oplysninger om Atlanterhavets colossale Blacksprutter. Forhandlinger ved de Skandinaviske Naturforskeres Syvende Mode, 7. 182-185.
- Sweeney, M. J. and C. F. E. Roper / N. A. Voss, M. Vecchione, R. B. Toll and M. J. Sweeney, eds., 1998: Classification, type localities and type repositories of recent Cephalopoda. Systematics and Biogeography of Cephalopods. Smithsonian Contributions to Zoology, 586 (I-II). 561-599.
- Winkelmann I., Campos P.F., Strugnell J., Cherel Y., Smith P.J., Kubodera T., Allcock L., Kampmann M.-L., Schroeder H., Guerra A., Norman M., Finn J., Ingrao D., Clarke M. & Gilbert T. P. (2013) Mitochondrial genome diversity and population structure of the giant squid <i>Architeuthis</i>: genetics sheds new light on one of the most enigmatic marine species. <i>Proceedings of the Royal Society, B</i> 280: 20130273.
 , 1860: Description de quelques fragments de deux Cephalopodes gigantesques. Verhandelingen der Koninklijke Akademie van Wetenschappen, Amsterdam, 9 (1). 16.
, 1860: Description de quelques fragments de deux Cephalopodes gigantesques. Verhandelingen der Koninklijke Akademie van Wetenschappen, Amsterdam, 9 (1). 16. K. and J. Knudsen, 1983: A catalogue of the type specimens of Cephalopoda (Mollusca) in the Zoological Museum, University of Copenhagen. Steenstrupia, 9 (10). 217-227.
K. and J. Knudsen, 1983: A catalogue of the type specimens of Cephalopoda (Mollusca) in the Zoological Museum, University of Copenhagen. Steenstrupia, 9 (10). 217-227. A. (1996). Molluscan name-bearing types in the Museum of New Zealand Te Papa Tongarewa. <em>Tuhinga: Records of the Museum of New Zealand Te Papa Tongarewa.</em> 9: 1-85.
A. (1996). Molluscan name-bearing types in the Museum of New Zealand Te Papa Tongarewa. <em>Tuhinga: Records of the Museum of New Zealand Te Papa Tongarewa.</em> 9: 1-85. 2. Rome, FAO. <em>pp. 121-123.</em>
2. Rome, FAO. <em>pp. 121-123.</em> , 1857: Oplysninger om Atlanterhavets colossale Blacksprutter. Forhandlinger ved de Skandinaviske Naturforskeres Syvende Mode, 7. 182-185.
, 1857: Oplysninger om Atlanterhavets colossale Blacksprutter. Forhandlinger ved de Skandinaviske Naturforskeres Syvende Mode, 7. 182-185. P. (2013) Mitochondrial genome diversity and population structure of the giant squid <i>Architeuthis</i>: genetics sheds new light on one of the most enigmatic marine species. <i>Proceedings of the Royal Society, B</i> 280: 20130273.
P. (2013) Mitochondrial genome diversity and population structure of the giant squid <i>Architeuthis</i>: genetics sheds new light on one of the most enigmatic marine species. <i>Proceedings of the Royal Society, B</i> 280: 20130273.Albums | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| none | Architeuthis Dux | Disease Ceremonial | Not On Label | none | US | 2012 | Продать эту версию | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| none | Architeuthis Dux | Not On Label | none | 2012 | Продать эту версию | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| InMe#006 | Architeuthis Dux | Inferior Meat Records | InMe#006 | US | 2013 | Продать эту версию | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| none | Architeuthis Dux | Not On Label | none | USA & Canada | 2013 | Продать эту версию | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| none | Architeuthis Dux | Not On Label | none | US | 2013 | Продать эту версию | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| none | Architeuthis Dux | Not On Label | none | USA & Canada | 2013 | Продать эту версию | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| none | Architeuthis Dux | Glossary Records | none | 2014 | Продать эту версию | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| #420-002 | Architeuthis Dux | #420Holocaust Records | #420-002 | US | 2014 | Продать эту версию | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| none | Balisticos Rutilantes, God Pussy, Architeuthis Dux | Not On Label | none | US | 2014 | Продать эту версию | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| none | Architeuthis Dux, Grandpa Lies Again | Inferior Meat Records | none | US | 2014 | Продать эту версию | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| none | Jeritan / Architeuthis Dux | Not On Label | none | Indonesia | 2016 | Продать эту версию | | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| infratape011 | Architeuthis Dux, Le TRASH CAN | Infra Ediciones | infratape011 | 2016 | Продать эту версию | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| none | Architeuthis Dux | No Part Of It | none | 2016 | Продать эту версию | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| none |
| Architeuthis Dux | No Part Of It | none | 2016 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Singles & EPs | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| none | Architeuthis Dux | Not On Label | none | USA & Canada | 2013 | Продать эту версию | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Miscellaneous | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| none | Architeuthis Dux | Not On Label | none | US | 2011 | Продать эту версию | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| REORH#055 | Architeuthis Dux, Tigre Merde | Ram Horn Records | REORH#055 | US | 2014 | Продать эту версию | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| KRE-31 | Architeuthis Dux / Arvo Zylo | Ka-Rye-Eye Tapes | KRE-31 | 2016 | Продать эту версию | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 credits a):not(.artist_in_title a)»>
credits a):not(.artist_in_title a)»> credits a):not(.artist_in_title a)»>
credits a):not(.artist_in_title a)»> credits a):not(.artist_in_title a)»>
credits a):not(.artist_in_title a)»> credits a):not(.artist_in_title a)»>
credits a):not(.artist_in_title a)»> credits a):not(.artist_in_title a)»>
credits a):not(.artist_in_title a)»> credits a):not(.artist_in_title a)»>
credits a):not(.artist_in_title a)»> credits a):not(.artist_in_title a)»>
credits a):not(.artist_in_title a)»>:max_bytes(150000):strip_icc()/giantsquid-noaa-56a5f7605f9b58b7d0df50da.jpg) credits a):not(.artist_in_title a)»>
credits a):not(.artist_in_title a)»> credits a):not(.artist_in_title a)»>
credits a):not(.artist_in_title a)»>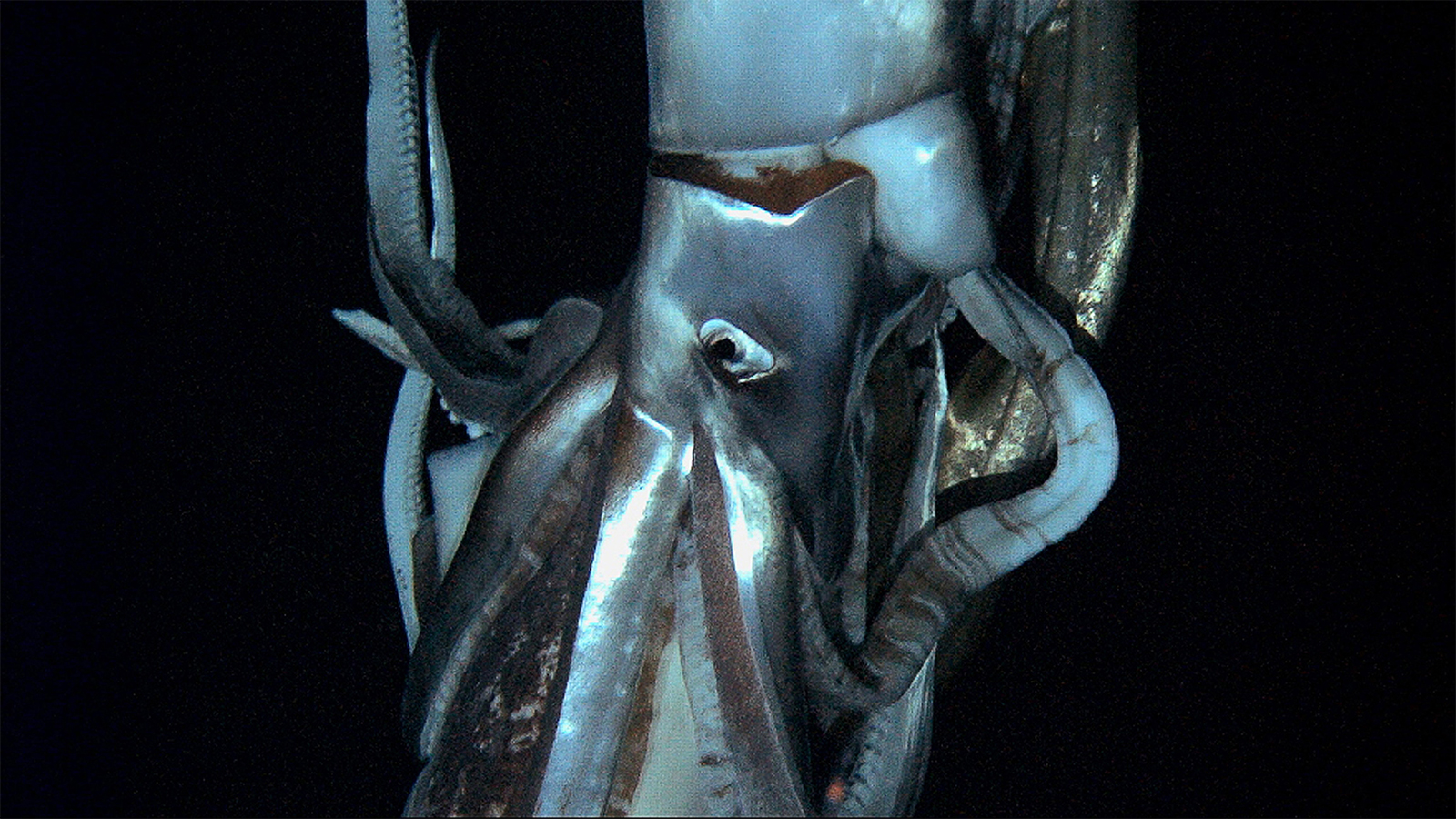 mr_toggler»>
mr_toggler»> credits a):not(.artist_in_title a)»>
credits a):not(.artist_in_title a)»> credits a):not(.artist_in_title a)»>
credits a):not(.artist_in_title a)»> credits a):not(.artist_in_title a)»>
credits a):not(.artist_in_title a)»> credits a):not(.artist_in_title a)»>
credits a):not(.artist_in_title a)»> credits a):not(.artist_in_title a)»>
credits a):not(.artist_in_title a)»> credits a):not(.artist_in_title a)»>
credits a):not(.artist_in_title a)»> credits a):not(.artist_in_title a)»>
credits a):not(.artist_in_title a)»>| Starting the trip | Начиная поездку |
| Waiting for it | Ждет его |
| Time has come, I can’t wait to face | Пришло время, я не могу дождаться встречи с тобой |
| The monster, it’s part of my fate, | Чудовище, это часть моей судьбы, |
| It’s my home | Это мой дом |
| Nom more sadness, only kindness lies in the ocean | Nom больше печали, только доброта лежит в океане |
| I found the beast and raise my fist ’cause I will not fail | Я нашел зверя и поднимаю кулак, потому что я не проиграю |
| Seeking for life through the waves | Ищу жизнь сквозь волны |
| I drift through the liquid world | Я плыву по жидкому миру |
| Making my own luck, I resign from your world | Мне повезло, я ухожу из твоего мира |
| Sealed in my Bathyscaph I agree | Запечатанный в моем батискафе, я согласен |
| Maybe there’s no safety when you come from the land | Может быть, нет безопасности, когда ты придешь с земли |
| But I want to ignore that huge place of terror | Но я хочу игнорировать это огромное место террора |
| Reach my incarnated fears | Достичь моих воплощенных страхов |
| Architeuthis, Architeuthis | Architeuthis, Architeuthis |
| Architeuthis Dux is mine | Architeuthis Dux — мой |
| Beholding its eyes | Созерцая его глаза |
| I stood still | Я остановился |
| Seeing … Reflections of a lost world | Видя . .. Отражения потерянного мира .. Отражения потерянного мира |
| I feel the experience life sized | Я чувствую опыт в натуральную величину |
| And the creature reveal it’s real | И существо показывает, что это реально |
| So I have kept my child side | Так что я сохранил свою детскую сторону |
| Yes I have kept my child side | Да, я сохранил свою детскую сторону |
Биологи получили редкие кадры гигантских глубоководных кальмаров Architeuthis dux
Исследователям удалось при помощи новаторской технологии глубоководной съемки запечатлеть представителей нескольких видов кальмаров, включая гигантского архитеутиса.
Гигантский кальмар архитеутис может вырастать до 14 метров в длину. Обнаружить этого моллюска достаточно сложно, поскольку он живет на глубине больше 400 м. У этих кальмаров развиты самые большие глаза, чтобы хорошо видеть в среде, куда почти не попадает солнечный свет. Ранее люди могли лишь видеть тела этих животных, выброшенные на берег, однако теперь ученым удалось заснять их под водой.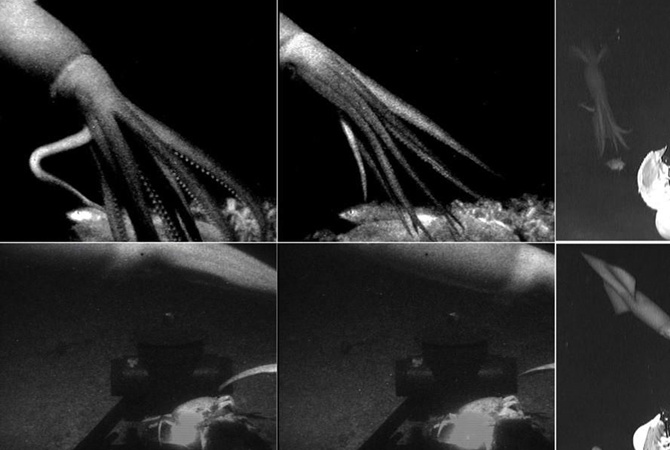
naked-science.ru
Команда биологов из Институт мыса Эльютера на Багамах, Университета Дьюка в Северной Каролине и Университета Южной Флориды (США) провела успешное исследование , результат которого опубликован в журнале PNAS.
Ученые рассказали, что для выполнения задачи использовали ненавязчивые платформы для глубоководных камер слабого освещения Medusa, оснащенные красными осветителями и приманками, которые имитируют биолюминесценцию. Такие устройства были развернуты в Большом Карибском бассейне, где и произошли встречи с крупными глубоководными кальмарами, включая гигантского кальмара Architeuthis dux, Pholidoteuthis adami, и двумя большими кальмарами — предположительно, Promachoteuthis. Эти виды обитали на глубине от 557 до 950 метров. По оценка биологов, длина мантии Promachoteuthis составляла порядка метра, Pholidoteuthis — 0,5 метра, а Archtiteuthis — 1,7 метра.
Ученые создали приманку E-Jelly, имитирующую биолюминесцентное изображение глубоководной медузы. Неоновый с синим экран E-Jelly давал понять кальмару, что неподалеку есть добыча. Из-за этого он подплывал ближе, в то время как камера его снимала.
Неоновый с синим экран E-Jelly давал понять кальмару, что неподалеку есть добыча. Из-за этого он подплывал ближе, в то время как камера его снимала.
naked-science.ru
Благодаря такому новаторскому методу ученым удастся детальнее изучить этот загадочный и редкий вид кальмаров.
«Каждая встреча с кальмарами дает новую информацию о ареале и распространении этих видов. Только три особи P. sloani нам удалось идентифицировать, и каждую из них обнаружили в районе Северо-Атлантического хребта. Если два вида, которых мы наблюдали, действительно были одним и тем же видом, это расширяет ареал P. sloani на 10 ° южной и западной широты до северной части Мексиканского залива», — говорят исследователи.
Ученые сняли на видео существо с огромными глазами, породившее легенды о Кракене
Натан Дж. Робинсон.
Гигантский кальмар Architeuthis dux до жути напугал моряков Древней Греции.
Океанологам и биологам США во главе с Эдит Виддер из Ассоциации исследований и охраны океана удалось запечатлеть гигантского кальмара Architeuthis dux. Этот тот самый вид, из-за которого моряки придумали легенду о Кракене. Он также лег в основу мифов Древней Греции об огромных морских животных, которые затягивают моряков и даже огромные корабли в морские глубины.
Гигантский кальмар Architeuthis dux – это крупнейшее беспозвоночное на планете. Может достигать 14 метров в длину, однако за ним не легко наблюдать, поскольку Architeuthis dux обитают на глубине более 400 метров. Туда почти не проникает солнечный свет. Это подчеркнули в исследовании, которое обнародовали в журнале Science Direct.
Выживая в темноте, кальмары так развили зрение, что стали обладателями самых больших глаз в животном мире: около 30 сантиметров в диаметре. Океанологам трудно фотографировать моллюсков на такой глубине. Обычно люди видели только мертвых «кракенов», когда те выбрасывались на берег.
— Обычные методы исследования морских глубин, включая использование сетей, пилотируемых подводных аппаратов и дистанционно управляемых устройств, в первую очередь подходят для изучения лежачих или медленно плавающих организмов, в то время как ловушки с наживкой, как правило, привлекают падальщиков, а не хищников, — пояснили ученые. — По этой причине для того, чтобы запечатлеть этот вид живых существ, пришлось разработать дискретные платформы для глубоководных камер. Использовались камеры, которые снимают при слабом освещении, красные огни и приманки, имитирующие биолюминесценцию.
A giant squid was caught on camera in 2019, emerging from darkness and grabbing at a lure attached to a deep sea research vessel. It was the first time one was filmed in the gulf of Mexico. The juvenile was estimated to be up to 3.7 meters long https://t.co/WJDljzNq55pic.twitter.com/BlzcKV9tqG
— Massimo (@Rainmaker1973) March 14, 2020
Команда биологов вооружилась устройством с очень тусклым красным светом, чтобы спрятать камеру от кальмаров. Эти моллюски не различают красный свет, отметили на Phys.org. Исследователи также использовали элемент под названием e-Jelly. Он имитирует свечение медузы рода Atolla с помощью синих неоновых огней. Это стало приманкой для моллюсков, которые охотятся на светящихся живых существ. Сочетание красных огней с приманкой e-Jelly позволило заснять кальмаров в японских и американских водах на глубине от 630 до 900 метров. Впервые этих моллюсков сняли в 2012 году, но на этот раз удалось сделать более подробный ролик.
Эти моллюски не различают красный свет, отметили на Phys.org. Исследователи также использовали элемент под названием e-Jelly. Он имитирует свечение медузы рода Atolla с помощью синих неоновых огней. Это стало приманкой для моллюсков, которые охотятся на светящихся живых существ. Сочетание красных огней с приманкой e-Jelly позволило заснять кальмаров в японских и американских водах на глубине от 630 до 900 метров. Впервые этих моллюсков сняли в 2012 году, но на этот раз удалось сделать более подробный ролик.
ЧИТАЙТЕ ТАКЖЕ
Сооснователь Neuralink заявил, что может создать Парк Юрского периода
Giant Squid (Architeuthis dux) · iNaturalist
- Explore
Community
- People
- Projects
- Journal Posts
- Forum
More
- Taxa Info
- Guides
- Places
- Site Stats
- Help
- Video Tutorials
- Managing Projects
- Teacher’s Guide
- Log In
or
Sign Up
Loading…
About
Help
Forum
Press
Our Blog
Community Guidelines
Terms of Service
Privacy
DONATE
STORE
Twitter
Instagram
Member of the iNaturalist Network
|
Powered by iNaturalist open source software
|
Documentation for developers
English
Toggle Dropdown
- العربية
- български
- Breton
- Català
- česky
- Dansk
- Deutsch
- Ελληνικά
- English
- Esperanto
- Español
- Español (Argentina)
- Spanish (Colombia)
- Español (México)
- Eesti
- Euskara
- suomi
- français
- Français (Canada)
- Galego
- עברית
- magyar
- bahasa Indonesia
- Italiano
- 日本語
- 한국어
- Lëtzebuergesch
- Lietuvių
- Latviešu
- македонски
- Norsk Bokmål
- Nederlands
- Occitan
- Polski
- Português
- Português (Brasil)
- Русский
- Slovenský
- Shqip
- Svenska
- Thai
- Türkçe
- Українська
- 简体中文
- 繁體中文
- Help Translate!
Эти морские твари такие гигантские, что их не может съесть кашалот
- Мэтт Уокер
- BBC Earth
Автор фото, SPL
Обозреватель
BBC Earth выяснил, что гигантские кальмары неспроста носят такое название – они действительно могут вырастать до невероятных размеров.
Рыбаки с незапамятных времен передавали из уст в уста истории о гигантском морском чудище кракене, которое выхватывало людей из лодок и тащило их в пучину океана, где их ждала гибель.
С тех пор как люди впервые увидели монстра, не стихают споры о его максимальном размере.
Теперь мы уже знаем, что кракен действительно существует: чудище из рыбацких рассказов — это вид гигантского кальмара.
И хотя маловероятно, чтобы кальмар мог убивать людей, он действительно бывает огромным — это одно из самых крупных известных науке беспозвоночных.
Но одного мы не знаем до сих пор: каких размеров может достигать гигантский кальмар?
Загадочные твари
Гигантский кальмар остается одним из самых загадочных крупных животных нашей планеты.
Записи о выброшенных на европейские берега останках особей, принадлежащих, судя по всему, к виду гигантских кальмаров Architeuthis dux (которые считаются самыми длинными), датируются далеким 1639 годом, однако гигантский кальмар в естественной среде обитания был впервые сфотографирован только в 2004 году.
Автор фото, Conrad Maufe naturepl.com
Подпись к фото,
Выброшенный на берег гигантский кальмар
«С тех пор, как люди впервые его увидели, не стихают споры о его максимальном размере», — рассказывает доктор биологических наук Крис Пакстон из Сент-Эндрюсского университета в Шотландии (Великобритания).
Пытаясь выяснить, какой длины могут достичь гигантские кальмары, он изучил останки особей A. dux; результаты этого анализа опубликованы в английском научном журнале Journal of Zoology, посвященном вопросам зоологии.
В рамках исследования он также рассмотрел исторические сведения и передаваемые из уст в уста рассказы очевидцев о гигантских кальмарах, за которыми люди наблюдали с кораблей, об останках, выброшенных на берег, и даже о частях тел этих монстров, найденных в желудках кашалотов (которые, как считается, охотятся на огромных головоногих и питаются ими).
В общей сложности тем или иным образом было измерено около 460 особей гигантского кальмара.
Захватывающее зрелище
Считается, что крупнейший из встречавшихся кальмаров достигал 53 метров в длину.
Это чудовище видели однажды ночью в годы Второй мировой войны недалеко от Мальдивских островов — его заметил человек по имени Дж. Д. Старки, находившийся на борту британского военного корабля.
Однако многие из подобных утверждений не следует принимать всерьез.
Автор фото, Christian Darkin Science Photo Library
Подпись к фото,
Останки кальмаров находят в брюхе у кашалотов
В рассказах очевидцев размер кальмаров зачастую сильно переоценивается. К примеру, как пишет в своей новой книге британский палеонтолог Даррен Наиш, Дж. Д. Старки утверждал, что виденное им чудовище было длиннее корабля, на котором он находился. И это без учета двух длинных щупалец кальмара, с помощью которых он питается и на которые обычно приходится более половины длины этого головоногого.
Измерение всего тела гигантского кальмара также может быть затруднительным, так как у извлеченного из воды монстра мягкие ткани могут стать эластичными и неестественным образом растянуться.
Поэтому Пакстон исключил из расчетов ненадежные сведения, а затем с помощью ряда методов оценил максимальный размер, которого может достичь гигантский кальмар.
Среди прочего он вычислил совокупную длину его тела путем экстраполяции размеров различных его частей, таких как клюв, твердая часть рта, используемая для кусания и поедания добычи, а также мантия – значительная часть туловища, расположенная прямо над головой и щупальцами моллюска.
Самые крупные особи
Как сообщил обозревателю BBC Earth Пакстон, длина мантии у самого крупного из достоверно измеренных кальмаров составила 2,79 метра.
«Этого кальмара выбросило на берег в мае 1879 года в проливе Кука (Новая Зеландия), — рассказывает ученый. — Однако его суммарная длина не была зафиксирована. Исходя из имеющихся данных, можно предположить, что она достигала 14,28 метра».
Впрочем, это не означает, что кальмар может достигать в длину максимум 14,28 метра, — по двум причинам.
Автор фото, fir0002 CC by NC
Подпись к фото,
Замороженный гигантский кальмар в Мельбурнском аквариуме (Австралия)
Во-первых, не у всех кальмаров части тела имеют одни и те же пропорции. Так, два кальмара с мантией одинакового размера могут иметь щупальца разной длины — точно так же, как у людей одного роста может различаться длина ног. А у кальмаров эти различия могут быть довольно большими.
Вторая причина состоит в том, что весьма маловероятно, чтобы среди пойманных и измеренных гигантских кальмаров непременно оказалась самая крупная из когда-либо живших на свете особей.
Так что вполне вероятно, что где-то существуют кальмары еще большего размера.
Судя по данным Пакстона, они могут быть очень и очень большими.
Самый длинный из всех
«Учитывая разнообразие особей в рамках популяции, при длине мантии 2,79 метра общая длина может варьироваться от 5,83 метра до 27,53 метра», — считает ученый.
Исходя из этого, можно предположить, что гигантские кальмары вырастают до 20 метров в длину и гипотеза о том, что некоторые из них могут достигать 27 и более метров, представляется «вполне правдоподобной».
«Однако как ответственный ученый я бы хотел подчеркнуть, что эти расчеты основаны на экстраполяции, — пояснил Пакстон. – И, как я говорю своим студентам, экстраполяция может оказаться ненадежным инструментом».
Впрочем, по его мнению, в этом случае экстраполировать данные можно довольно уверенно, и даже если другие специалисты по гигантским кальмарам не согласны с полученными результатами, он надеется, что в будущем их можно будет проверить и подтвердить благодаря новым находкам.
Кроме того, исходя из данных этого исследования, напрашивается еще один вывод.
Подводные битвы
В легендах и мифах о морских чудовищах упоминаются подводные «битвы титанов» — гигантских кальмаров и кашалотов.
Однако некоторые кальмары могут вырастать до таких размеров, что кашалоты просто не смогут их поймать.
Автор фото, Mark Carwardine naturepl.com
Подпись к фото,
Кашалот пожирает добычу — возможно, гигантского кальмара
Судя по данным анализа Пакстона, возможно, самый крупный кальмар имеет слишком большую длину, чтобы его могла поймать самка кашалота, которая вырастает до 11-12 метров в длину.
За такими крупными особями способны охотиться разве что самые большие самцы кашалотов, которые достигают в длину 20 метров.
ADW: Architeuthis dux: ИНФОРМАЦИЯ
Географический ареал
Гигантские кальмары распространены во всех океанах мира, обычно на континентальных и островных склонах. Концентрации найденных видов находятся в северной части Атлантического океана, особенно в Ньюфаундленде, Норвегии, северных Британских островах и океанических островах Азорские острова и Мадейра; Южная Атлантика в водах юга Африки; северная часть Тихого океана вокруг Японии и юго-западная часть Тихого океана вокруг Новой Зеландии и Австралии; циркумглобальный в Южном океане.Редко встречаются из тропических и высоких полярных широт (Forch 1998)
.
Среда обитания
Никто на самом деле не знает, где живут гигантские кальмары, потому что никто не видел ни одного живого в естественной среде обитания. Только недавние исследования показали, где может быть эта среда обитания. Он находится в глубоком море, возможно, на глубине от 200 до 1000 метров, и, возможно, он связан с дном моря, а не в середине воды. С другой стороны, особи, пойманные в сети, иногда происходят из середины воды.
Работа, проделанная доктором Оле Бриксом из Бергенского университета, показала, что кровь кальмаров не очень хорошо переносит кислород при более высоких температурах. Кальмар задохнется в теплой воде. Теплая вода заставит гигантского кальмара подняться на поверхность и не сможет вернуться обратно. Так что гигантских кальмаров, скорее всего, можно будет найти в более прохладной воде. (Форч, 1998 г.) (Банистер и Кэмпбелл, 1985 г.)
Физическое описание
Architeuthidae — самые большие известные головоногие моллюски, самые крупные из известных моллюсков и, вероятно, самые крупные беспозвоночные, когда-либо существовавшие в океанах. Architeuthidae имеют общую длину 60 футов, большинство обнаруженных экземпляров действительно находятся в диапазоне 35-45 футов. Есть еще много таких, длина которых колеблется от 20 до 30 футов. Общая длина включает тело, голову, руки и два длинных кормящих щупальца. Эти кормящие щупальца намного длиннее остального тела. Самые тяжелые животные весят около тонны, но в большинстве случаев их вес составляет тысячу фунтов или меньше.
У этих гигантских кальмаров самые большие глаза среди всех животных в мире.Глаза гигантского кальмара могут быть размером с человеческую голову. У большинства глубоководных животных очень большие глаза, поэтому они могут собирать небольшое количество света, доступного в глубоких глубинах океана. Они могут даже видеть биолюминесцентный свет.
У Architeuthidae есть два щупальца, которые в среднем составляют около 10-12 метров в длину. У этих щупалец есть много присосок на концах, называемых булавами. Щупальца узкие и имеют присоски, которые представляют собой субсферические чашечки, выстланные острыми, мелко зазубренными кольцами хитина, расположенными в четыре продольных ряда.Эти присоски покрывают только внутреннюю поверхность рук и щупалец. Эти щупальца делятся на запястье, кисть и дактиль. Кисть имеет увеличенные присоски вдоль двух медиальных рядов. Неизвестно, что присоски на щупальцах и руках больше пяти-пяти с половиной сантиметров. В области запястья густое скопление присосок в шесть-семь неправильных поперечных рядов.
У Architeuthidae также есть пропорционально маленькие, яйцевидные плавники без свободных передних долей.Плавники в задней части мантии используются, чтобы помочь кальмарам двигаться мягкими ритмичными импульсами воды, выталкиваемой из полости мантии по всей воронке.
Еще у них восемь рукавов с присосками в два продольных ряда. На концах рук у них есть клювы, похожие на попугайские клювы у основания. Еще одна особенность кальмаров заключается в том, что у них есть соединительные элементы со щек, которые прикрепляются к спинным краям рук.
Гигантский кальмар содержит темные чернила цвета сепии, которые у нас ассоциируются с более мелкими и более знакомыми кальмарами.
У них есть две очень большие жабры, покоящиеся внутри мантии. Кальмары способны дышать и быстро двигаться, расширяя полость мантии за счет сокращения наборов мускулов внутри мантии. Вода заполняет расширенное пространство, мышцы расслабляются, а эластичная мантия возвращается к меньшему размеру, выбрасывая воду через воронку. Струя воды закрывает створки по обе стороны головы кальмара, поэтому вода может выходить только через воронку.
Нервная система кальмаров очень обширна, и у них даже есть сложный мозг.По этой причине они подвергаются обширным исследованиям. Система кровообращения закрыта, что является отличительной чертой кальмаров (Portner, et al., 1994) (Forch, 1998),
.
Репродукция
Репродукция Architeuthis малоизвестна. Гипотезы основаны на наблюдениях за половыми признаками мертвого Architeuthis и на знаниях других кальмаров
.
Самки производят огромное количество яиц от беловатого до кремового цвета, около.5–1,4 мм в длину и 0,3–7 мм в ширину, в зависимости от стадии созревания. У одной самки в яичнике было более 5000 г (более 11 фунтов) яиц, что намного превышает миллион яиц. Как и у большинства эгопсид, самки имеют единственный срединный яичник в заднем конце мантийной полости, парные извитые яйцеводы вместе со зрелыми яйцами проходят, затем выходят через яйцевидные железы и большие нидаментальные железы, которые продуцируют количество студенистого материала. Неизвестно, откладываются ли яйца в большую студенистую матрицу, как у большинства крупных океанических кальмаров, или выпускаются по отдельности, хотя большие нидаментальные железы предполагают первый метод.
Самцы, как правило, достигают половой зрелости меньшего размера, чем самки. Сообщается, что два вентральных плеча модифицируются для передачи сперматофоров самке. Как и у большинства других головоногих моллюсков, один задний семенник производит сперму, которая перемещается в сложную систему желез, производящих сперматофоры. Они хранятся в продолговатом мешочке или мешочке Нидхема, из которого они выбрасываются во время спаривания. Сумка Нидхема полностью зрелых самцов заполнена сотнями сперматофоров.Мешочек Нидхема оканчивается половым членом. Пенис настолько удлинен, что выходит вперед за отверстие мантии. Хотя спаривание не наблюдалось и точная роль полового члена неясна, у некоторых женщин были обнаружены сперматангию, содержащие сперму мешочки сперматофора, встроенные в ткань вокруг оснований рук и головы.
Головоногие моллюски известны как очень быстрорастущие животные. Некоторые виды мелких мелководных форм достигают половой зрелости через 6-8 месяцев, а большинство видов, по которым имеются данные о росте, возрасте и зрелости, достигают репродуктивной способности в течение 12-18 месяцев.Многие из обнаруженных экземпляров Architeuthis были зрелыми, особенно самки. Но возраст зрелости Architeuthis точно неизвестен. Одно исследование предполагает, что взрослый размер достигается в течение 3 лет. Даже при ожидаемых быстрых темпах роста головоногих моллюсков достижение массы в 500 кг или более менее чем за 3 года является впечатляющим.
(Несис 1987)
Поведение
Поскольку ученые никогда не наблюдали гигантских кальмаров живыми в их естественной среде обитания, они не могут сказать, на что похоже его поведение.Невозможно точно узнать, как пользователи Architeuthis взаимодействуют друг с другом. Ученые выдвинули гипотезу о том, что Architeuthis могут быть одиночными охотниками. Они верят в это, потому что никакие два Architeuthis никогда не попадались в рыболовные сети вместе. (Forch 1998)
Привычки питания
В течение многих лет никто не знал, чем питаются гигантские кальмары. Это потому, что они никогда не наблюдались в дикой природе.Некоторые недавние исследования мертвых людей показали, что гигантские кальмары едят глубоководных рыб, таких как апельсиновый ерш и хоки. Они также едят другие виды глубоководных кальмаров, но не гигантских кальмаров Architeuthis .
Какими бы большими ни были эти животные, они, вероятно, смогли бы поймать почти все, может быть, даже китов (см. Комментарии)! Они захватывают свою добычу двумя длинными щупальцами для кормления. Щупальца выстреливаются, чтобы схватить добычу. Присоски на концах щупалец хватают добычу, и щупальца сжимаются, перенося добычу в руки.Затем руки еще больше подчиняют добычу, притягивая ее к сильным острым клювам. Клювы управляются массивным набором мускулов, которые позволяют им прокусывать практически все, что может захватить кальмар. Но кусочки пищи гигантского кальмара перед перевариванием необходимо измельчить. На языке есть орган, известный как радула, в котором расположены ряды маленьких зубов, похожих на напильники. Радула измельчает пищу еще до того, как язык проталкивает ее по пищеводу к органам пищеварения.
Хищников взрослых гигантских кальмаров очень мало. Самый лучший и, наверное, единственный — кашалот. В младенчестве и молоди у них много педаторов, в основном глубоководных рыб. Как только гигантские кальмары превышают определенный размер, они перерастают размер большинства своих потенциальных хищников. Кашалоты вырастают до 40-50 футов в длину, но весят 30-40 тонн. Таким образом, даже несмотря на то, что гигантский кальмар огромен, он недостаточно велик, чтобы убежать или сразиться с кашалотом. В большинстве случаев побеждает кашалот.Об этом свидетельствует количество гигантских кальмаров, обнаруженных в желудке кашалота. (Форч, 1998 г.) (Банистер и Кэмпбелл, 1985 г.)
Экономическое значение для людей: положительный результат
Поскольку известно немногое, трудно сказать, насколько это важно для людей. Это может быть неотъемлемая часть пищевой цепи, и если она будет нарушена, то может нанести вред китам, для чего у нас действительно есть применение. У кальмаров и других головоногих моллюсков очень отчетливая и сложная нервная система и мозг.Гигантский кальмар может помочь нам понять и узнать больше о нервной системе, возможно, даже о нашей. (Несис 1987, Гилберт и др. 1990)
Экономическое значение для людей: отрицательно
На данный момент никаких вредных воздействий на человека не наблюдается. Мы только недавно узнали о его существовании! Единственная незначительная проблема, которую он представляет, заключается в том, что он может запутаться в рыболовных сетях, но это не является серьезной проблемой. (Несис 1987)
Статус сохранения
Число особей этого вида неизвестно.(Forch 1998)
Поскольку немногие из этих видов были замечены живыми, существует множество историй, мифов и загадок о гигантских кальмарах, и часто они связаны с опасностью. Гигантского кальмара даже назвали морским чудовищем. Новые технологии, вероятно, позволят нам не только искать и, надеюсь, найти гигантского кальмара, но и исследовать и открывать для себя глубоководные места обитания.
Авторы
Джеррод Воан (автор), Юго-Западный университет, Стефани Фабрициус (редактор), Юго-Западный университет.
Глоссарий
- Северный Ледовитый океан
— водоем между Европой, Азией и Северной Америкой, расположенный в основном к северу от полярного круга.
- Атлантический океан
— водоем между Африкой, Европой, южным океаном (выше 60 градусов южной широты) и западным полушарием.Это второй по величине океан в мире после Тихого океана.
- Тихий океан
водоем между южным океаном (выше 60 градусов южной широты), Австралией, Азией и западным полушарием. Это самый большой океан в мире, покрывающий около 28% поверхности мира.
- придонный
Обозначает животное, которое живет на дне водоема или рядом с ним. Также водный биом, состоящий из дна океана ниже пелагических и прибрежных зон. Донные места обитания в самых глубоких океанах (ниже 9000 м) иногда называют абиссальной зоной.см. также океанический выход.
- двусторонняя симметрия
, имеющий такую симметрию тела, что животное можно разделить в одной плоскости на две зеркальные половины. У животных с двусторонней симметрией есть спинная и вентральная стороны, а также передний и задний концы. Синапоморфия билатериев.
- экзотермический
животных, которые должны использовать тепло, полученное из окружающей среды, и поведенческие адаптации для регулирования температуры тела
- родной диапазон
район, в котором животное обитает в природе, регион, в котором оно является эндемиком.
Список литературы
Банистер, К., А. Кэмпбелл. 1985. Энциклопедия водной жизни. Нью-Йорк, штат Нью-Йорк: факты в файле.
Forch, E. 1998. Морская фауна Новой Зеландии: головоногие моллюски: Oegopsida: Architeuthidae (гигантские кальмары). Веллингтон: Национальный институт водных и атмосферных исследований.
Д. Гилберт, У. Адельман, Дж. Арнольд. 1990 г.Кальмары как экспериментальные животные. Нью-Йорк, штат Нью-Йорк: Plenum Press.
Несис, К. 1987. Головоногие моллюски мира: кальмары, каракатицы, осьминоги и их союзники. Нептун, Нью-Джерси: Публикации T.F.H.
Портнер, Х., Р. О’Дор, Д. Макмиллан. 1994. Физиология головоногих моллюсков. Швейцария: издательство Gordon and Breach.
Гигантский кальмар | Смитсоновский океан
Разнообразие кальмаров
Кальмары бывают самых разных размеров, от меньших, чем ваш большой палец, до огромных гигантских кальмаров.(Ким Меллер / Смитсоновский институт)
Гигантский кальмар — это не просто один вид — или это так? Некоторые исследователи считают, что в роду Architeuthis (по-гречески «главный кальмар») насчитывается до 8 видов, каждый из которых представляет собой особый вид гигантских кальмаров. Но другие исследователи считают, что в мировом океане плавает только один Architeuthis . Единого мнения нет, потому что кальмаров очень сложно отследить, а доступных для изучения экземпляров так мало.
Однако очевидно, что Architeuthis имеет множество эволюционных родственников.В океане обитает около 500 видов кальмаров, и почти все они относятся к тому же таксономическому порядку, что и гигантский кальмар, называемый эгопсиной. Некоторые из них на удивление крошечные — всего около 2,5 сантиметра в длину. Другие впечатляюще велики, в том числе колоссальный кальмар ( Mesonychoteuthis hamiltoni ), который может вырасти даже больше, чем гигантский кальмар, достигая 45 футов (14 метров).
Эти виды кальмаров тесно связаны с улитками, моллюсками и даже слизнями: все они моллюски, которые определяются их мягким телом.Некоторые из этих мягких тел заключены в твердые оболочки, такие как моллюски и улитки, но не кальмары.
Кальмары принадлежат к особенно успешной группе моллюсков, называемых головоногими моллюсками, существующим около 500 миллионов лет назад. У некоторых предков современных кальмаров были раковины, такие как аммониты, которые управляли волнами 400–65 миллионов лет назад. Из тех, что еще существуют, одна небольшая группа — наутилусы — имеет внешнюю оболочку. Другой — который включает кальмаров, каракатиц и осьминогов — нет, хотя у кальмаров и каракатиц есть внутренняя опора, напоминающая позвоночник, сделанная из хитина, называемая пером.
Панцирь или без панциря, все головоногие моллюски имеют хорошо развитый мозг и очень активны, стремительно перемещаясь через океан. У большинства есть чернильные мешочки. И многие могут изменить цвет и текстуру кожи в мгновение ока.
Гигантский кальмар | National Geographic
- Общее название:
- Giant Squid
- Научное название:
- Architeuthis dux
- Тип:
- Беспозвоночные
- Диета:
- Плотоядные
- Школа
- Размер:
- 33 фута
- Вес:
- 440 фунтов
Группа
- Статус Красного списка МСОП:?
- Наименее опасные
Наименее опасные Вымершие
- Текущая динамика численности населения:
- Неизвестно
Гигантский кальмар остается в значительной степени загадкой для ученых, несмотря на то, что он является самым большим беспозвоночным на Земле.Самый большой из этих неуловимых гигантов, когда-либо найденных, имел длину 59 футов и весил почти тонну.
Неуязвимость
Тем не менее, их негостеприимная глубоководная среда обитания делает их уникально трудными для изучения, и почти все, что ученые знают о них, основано на трупах, выброшенных на берег или вытащенных рыбаками. Однако в последнее время судьба ученых, изучающих этих неуловимых существ, начала меняться. В 2004 году японские исследователи сделали первые снимки живого гигантского кальмара.А в конце 2006 года ученые из Национального музея науки Японии поймали и вытащили на поверхность живую самку гигантского кальмара высотой 24 фута.
Массивные глаза
Гигантский кальмар и его двоюродный брат, колоссальный кальмар, имеют самые большие глаза в животном мире, их диаметр составляет около 10 дюймов. Эти массивные органы позволяют им обнаруживать объекты в темных глубинах, где большинство других животных ничего не увидят.
Диета
Как и у других видов кальмаров, у них есть восемь рук и два более длинных кормящих щупальца, которые помогают им подносить пищу к своим клювообразным ртам.Их диета, вероятно, состоит из рыбы, креветок и других кальмаров, а некоторые предполагают, что они могут даже атаковать и съесть маленьких китов.
Locomotion
Они маневрируют своим массивным телом с плавниками, которые кажутся крошечными для их размера. Они используют свою воронку в качестве двигательной установки, втягивая воду в мантию или основную часть тела и вытесняя ее назад.
Диапазон
Ученые недостаточно знают об этих животных, чтобы точно сказать, каков их ареал, но туши гигантских кальмаров были обнаружены во всех океанах мира.
Особенности жизненного цикла гигантского кальмара Architeuthis dux, выявленные с помощью сигнатур стабильных изотопов, записанных в клювах | Журнал ICES по морским наукам
Аннотация
Герра, А., Родригес-Наварро, А. Б., Гонсалес, А. Ф., Романек, К. С., Альварес-Льорет, П. и Пирс, Г. Дж. 2010. Особенности жизненного цикла гигантского кальмара Architeuthis dux , выявленные по сигнатурам стабильных изотопов, записанным в клювах. — Журнал морских наук ICES, 67: 1425–1431.
Профили изотопов углерода и азота, построенные из верхних клювов четырех гигантских кальмаров Architeuthis dux из Бискайского залива и вод Намибии, позволили получить интегрированные во времени записи об их рационе. Значения δ 15 N варьировали от 5,5 до 13,4 ‰, а δ 13 C — от −14,4 до −17,8. Профили изотопов азота значительно различались по форме у четырех проанализированных животных, но δ 15 N увеличивалось вдоль каждого профиля с самыми низкими значениями вокруг кончика рострального кончика.Разница между самым низким и самым высоким значениями δ 15 N составила ∼5,8 ‰, что немного меньше двухуровневой разницы между трофическим положением малого и большого A. dux . Увеличение значений δ 13 C было не столь выраженным, как для профилей δ 15 N, но изменения предполагают онтогенетический сдвиг в рационе питания в раннем возрасте от более мелкой добычи с относительно низким трофическим статусом к более крупной жертве с более высоким статусом. Колебания значений δ 13 C, наблюдаемые около верхушки рострального конуса, могут быть связаны с большей внутренней изменчивостью изотопного состава углерода относительно небольшой добычи и / или временным миграционным поведением в раннем возрасте.Относительная стабильность профилей δ 13 C на протяжении оставшейся части их жизни согласуется с гипотезой о том, что взрослые гигантские кальмары населяют относительно небольшие, четко очерченные и продуктивные районы, где пищевые ресурсы имеют постоянный изотопный состав углерода.
Введение
Традиционно исследования рациона головоногих моллюсков основывались на анализе содержимого их желудков (Nixon, 1987; Rodhouse, Nigmatullin, 1996). Однако этот прямой метод часто дает только частичные и необъективные результаты (Boyle and Rodhouse, 2005).Одна из проблем — сложность или невозможность получения информации о долгосрочных привычках кормления (Jackson et al ., 2007). В настоящее время, по крайней мере, два непрямых метода предлагают такую временную интеграцию, анализ жирных кислот и стабильных изотопов тканей головоногих (Stowasser et al ., 2006; Jackson et al ., 2007).
Соотношения встречающихся в природе стабильных изотопов азота ( 15 N / 14 N, δ 15 N) и углерода ( 13 C / 12 C, δ 13 C) в обоих твердых мягкие ткани у разных видов различаются.Ткани потребителей обогащены δ 15 N примерно на 3–5 ‰ по сравнению с их пищей, что обеспечивает механизм для оценки трофического уровня (Hobson, 1990; McCutchan et al. ., 2003; Vanderklift and Ponsard, 2003; Bode et al ., 2004; Cherel et al ., 2009a). Напротив, δ 13 C обычно показывает увеличение только на ~ 1 ‰ на трофический уровень, а соотношение изотопов углерода обычно интерпретируется как предоставление информации о происхождении первичных источников органического вещества в трофической сети.В морской среде значения δ 13 C можно использовать для различения вкладов низких и высоких широт, прибрежных и морских, пелагических и бентических вкладов в потребление пищи (Rau et al. ., 1982; Hobson et al ., 1994; Smith et al. ., 1996; McCutchan et al. ., 2003; Cherel and Hobson, 2007).
Клювы кальмаров — это твердые структуры, состоящие из хитин-белковых комплексов (Hunt and Nixon, 1981), которые сопротивляются перевариванию и могут накапливаться в желудках хищников (Clarke, 1980).Из-за постепенного увеличения инертного биологического материала они сохраняют молекулярную запись роста на протяжении всего жизненного цикла, которую можно расшифровать, чтобы лучше понять трофическую экологию этих животных (Cherel and Hobson, 2005).
Несколько исследований подчеркнули потенциал стабильных изотопов для изучения трофических уровней головоногих моллюсков (Takai et al ., 2000; Ruiz-Cooley et al ., 2004; Cherel and Hobson, 2005). Эксперименты с контролируемым кормлением показали, что сигнатуры стабильных изотопов мышц головоногих моллюсков можно использовать для различения животных, питающихся разными диетами, как в исследованиях в неволе (Hobson and Cherel, 2006; Stowasser et al ., 2006) и сравнение содержимого желудка (Ruiz-Cooley et al ., 2006). Изотопные вариации между и внутри тканей у диких океанических кальмаров Todarodes filippovae также предполагают изменение рациона с более низких трофических уровней на более высокие во время роста (Cherel et al. Парри (2008) использовал различия в соотношении изотопов азота, чтобы продемонстрировать, что взрослые особи Ommastrephes bartramii находятся на один трофический уровень выше, чем взрослые особи Sthenoteuthis oualaniensis . Cherel и др. .(2009b) обнаружили, что изотопные сигнатуры азота и углерода являются полезными инструментами для документирования трофической структуры сообщества глубоководных головоногих моллюсков, включая гигантского кальмара.
Дигл и др. . (2005) исследовали использование методов полимеразной цепной реакции (ПЦР) для обнаружения ДНК в останках жертв из образца кишечника Architeuthis . Амплифицированная область ДНК различалась по размеру, что позволяло разделить компоненты рыб и кальмаров. Рыба была идентифицирована как Macruronus novaezelandiae , также был обнаружен каннибализм.Однако наблюдалась высокая частота ложных аллелей, генерируемых ПЦР, и это может быть проблематичным при скрининге видов жертв, представленных низкой частотой ДНК.
Относительно недавним нововведением в исследованиях стабильных изотопов стало использование регистрирующих структур, таких как зубы китообразных, усатый ус и статолиты головоногих, для отслеживания изменений изотопных сигнатур в течение жизни отдельного животного. Поскольку изотопный состав каждого слоя роста будет отражать местонахождение животного и его пищевые привычки во время его осаждения, такую информацию можно интерпретировать с точки зрения онтогенетической изменчивости рациона и моделей миграции (Boyle and Rodhouse, 2005; Field et al. al ., 2007; Mendes et al ., 2007).
Цель исследования — понять долгосрочные пищевые привычки и трофическое положение Architeuthis dux с помощью анализа изменений в составе стабильных изотопов вдоль верхней части клюва. Мы предполагаем, что, поскольку передний конец рострума представляет ранние стадии жизни животного, а задний край капюшона — самую последнюю стадию, реконструкция индивидуальных трофических историй должна быть достижимой. Это первое исследование, в котором клювы головоногих используются подобным образом.
Материалы и методы
Мы проанализировали трех созревающих самок A. dux (таблица 1), которые были выброшены на мель на пляже Колунга (43 ° 30′06.36′′N 5 ° 15′39.41′′E; Бискайский залив) в 2001 и 2003 годах, и одна Неполовозрелая самка вылавливалась в 1990 г. траулером выдры, работавшим на глубине 350–390 м в водах Намибии (таблица 1). После сбора созревающие животные были заморожены при -20 ° C. Незрелый образец замораживали на борту при -40 ° C. Клювы удаляли у размороженных животных в лаборатории при комнатной температуре, очищали с использованием раствора 5% соляной кислоты пепсина в дистиллированной воде (pH 2) для удаления поверхностных остатков мягких тканей, затем консервировали в 70% этаноле.Мы выбрали верхний клюв, потому что несколько авторов использовали верхний клюв для определения возраста (Raya and Hernández-González, 1998; Hernández-López et al ., 2001; Higgins et al ., 2009; Perales-Raya et al. ., 2009).
Таблица 1.
Анализируемые образцы Architeuthis .
Таблица 1.
Проанализированы образцы Architeuthis .
Методология стабильных изотопов
Для изотопного анализа квадраты размером ∼1 мм × 1 мм были вырезаны из верхней части клюва каждого образца (рис. 1) каждые 1.От верхушки рострального до заднего края 43 мм. Подвыборки (1 мг) загружали в предварительно очищенные оловянные чашки и помещали в элементный анализатор (NC 2500, CE Instruments), присоединенный к масс-спектрометру с непрерывным потоком изотопов (Delta plus XL, Finnigan) для анализа стабильных изотопов углерода и азота. Отношения стабильных изотопов ( 13 C / 12 C и 15 N / 14 N) представлены в единицах на тысячу (‰) с использованием δ-обозначения по отношению к белемниту Vienna Pee Dee (углерод). Международный стандарт.Международный сертифицированный стандарт (DORM-2) периодически анализировался, и результаты были воспроизводимы с точностью выше ± 0,1 ‰ [1σ (стандартное отклонение), n = 38] для значений δ 13 C и δ 15 N. Мы также рассчитали соотношение C / N в каждом клюве.
Рис. 1.
Верхний клюв Architeuthis . УХЛ, длина верхнего капюшона (по Кларку, 1980).
Рис. 1.
Верхний клюв Architeuthis . УХЛ, длина верхнего капюшона (по Кларку, 1980).
Статистика
Поскольку материал для изотопного анализа был получен примерно через каждые 1,5 мм на длине 70 мм капюшона самого большого верхнего клюва, было всего 47 отдельных сегментов, а другая серия была короче. Все наборы данных были протестированы на автокорреляцию. Автокорреляция наблюдалась для лагов 1–3 позиций, но частичные автокорреляции обычно были значительными ( p <0,05) только для лага 1 (данные по азоту) или лагов 1 и 2 (данные по углероду).
Длина наборов данных различалась между животными, и по этой причине мы аппроксимировали авторегрессионную структуру порядка 1 или 2 [AR (1) или AR (2)], включив изотопные отношения для предыдущих одного или двух сегментов как линейные. предикторы. При использовании этого подхода изотопные отношения для первых одного или двух сегментов в каждой серии (начиная с кончика рострального слоя) исключаются из анализа, поскольку предыдущие значения не определены. Обобщенные аддитивные модели (GAM) были подогнаны к данным отношения изотопов, причем отношение изотопов было переменной отклика.Положение (номер сегмента) рассматривалось как непрерывная независимая переменная (аппроксимированная гладкой кривой), а животное использовалось как категориальная независимая переменная. Значения изотопов были приблизительно нормально распределены, поэтому использовалась GAM по Гауссу с функцией идентичности связи. Мы проверили, различаются ли онтогенетические тенденции (то есть влияние положения) у разных животных. Следовательно, полная модель для азота была
Полная модель для углерода включает дополнительный член для влияния предпоследнего изотопного отношения.Для обоих изотопов полная модель сравнивалась с использованием теста F со следующими упрощенными моделями: (i) модель с отдельными сглаживателями для эффекта положения для каждой страны, с использованием страны (а не отдельного лица) в качестве категориального критерия. объясняющая переменная; (ii) модель с комбинированным сглаживателем для всех животных для эффекта положения; (iii) модель без предыдущих соотношений изотопов в качестве предикторов. Всегда проверялись остатки модели на наличие закономерностей. Все модели были подогнаны с использованием программного обеспечения BRODGAR (www.brodgar.com), интерфейс для языка статистического программирования R, и процедуры были основаны на описанных в Mendes et al . (2007) и Зуур и др. . (2007).
Результаты
Значения δ 15 N в капюшоне верхней части клюва у четырех образцов Architeuthis , использованных в этом исследовании (см. Приложение с исходными данными), варьировались от 5,5 до 13,4 ‰ (Рисунок 2). Обогащение δ 15 N, наблюдаемое между верхушкой рострального конца и задним краем капюшона, варьировалось в пределах 5.2 и 7,4 (в среднем 5,8). Принимая во внимание, что ростральный кончик представляет собой самый ранний этап жизни, а задний край капюшона — взрослую стадию жизни, наблюдалась общая тенденция к увеличению обогащения δ 15 N с течением времени, что согласуется с онтогенетическим увеличением трофической уровень. Изменение наклона кривой свидетельствует о том, что скорость повышения трофического уровня выше в раннем периоде жизни (как зарегистрировано ближе к верхушке рострального канала; см. Рис. 2), хотя следует иметь в виду, что такая интерпретация неверна. на основе скорости изменения соотношений изотопов по отношению к единицам роста клюва, а не единицам времени.
Рис. 2.
Профили изотопов азота вдоль капюшона верхнего клюва самки Architeuthis . Номера образцов как в таблице 1. Расстояние (в мм) вдоль бленды от кончика рострального отверстия.
Рис. 2.
Профили изотопа азота вдоль капюшона верхнего клюва самки Architeuthis . Номера образцов как в таблице 1. Расстояние (в мм) по капюшону от кончика рострального кончика.
Значения δ 13 C находились в диапазоне -14.От 3 до -17,8 ‰. Они увеличились с -17,8 до -15,8 от вершины рострума до заднего края капюшона у испанских экземпляров (1932, 1964 и 1963 гг.) И с -16,8 до -14,3 у намибийских (1427). Примечательно, что наблюдались некоторые колебания в значениях δ 13 C вблизи кончика рострального кончика, и также кажется, что соотношение изотопов достигает пика, а затем начинает уменьшаться к концу жизни животного (Рисунок 3). Отношение масс C / N варьировало от 1,99 ± 1,25 до 1,42 ± 0,35, уменьшаясь от рострума к краю верхнего капюшона клюва (Приложение).
Рис. 3.
Профили изотопов углерода вдоль капюшона верхнего клюва самок Architeuthis . Номера образцов как в таблице 1. Расстояние (в мм) по капюшону от кончика рострального кончика.
Рис. 3.
Профили изотопов углерода вдоль капюшона верхнего клюва самок Architeuthis . Номера образцов как в таблице 1. Расстояние (в мм) по капюшону от кончика рострального кончика.
Полный GAM как для изотопов азота, так и для изотопов углерода показал лучшие результаты, чем любая из более простых моделей (см. Таблицы 2 и 3 для получения подробной информации о лучших моделях), а в остатках не было важных закономерностей.Следовательно, модель, которая позволяет каждому человеку иметь различную онтогенетическую траекторию, значительно лучше, чем модель, в которой предполагается, что все животные имеют одинаковые онтогенетические тенденции, или модель, в которой разрешены разные тенденции для Испании и Намибии. Тем не менее, как видно на рисунках 2 и 3, клюв намибийского животного был больше обогащен как C, так и N, чем любое из трех испанских животных. Подходящие сглаживания для эффекта положения обычно были нелинейными, о чем свидетельствуют степени свободы для сглаживателей, обычно превышающие 1.0 (кроме азота у животных 1964 г.). Как и ожидалось при просмотре необработанных данных (рисунки 2 и 3), все сглаживающие устройства (не показаны) показали в целом тенденцию к повышению в соотношении изотопов C и N по мере того, как положение вдоль клюва перемещалось дальше от кончика рострального кончика.
Таблица 2. Результаты GAM
для соотношений изотопов азота в вытяжке ( n = 163, скорректировано r 2 = 0,983).
| Параметр . | edf . | Коэффициент . | ST . | Значение . | Вероятность . | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Предыдущее соотношение | — | 0,510 | т | 13,50 | <0,0001 | |||||||
| Животное 1932 | — | −1,025 | — | −1,025 | — | −1,025 <0,0001 | ||||||
| Животное 1963 | — | −0.788 | т | −11,32 | <0,0001 | |||||||
| Животное 1964 | — | −0,851 | т | −11,57 | −11,57 | — | F | 15,73 | <0,0001 | |||
| Положение: животное 1932 | S (4,88) | — | 905 01 | <0,0001 | ||||||||
| Позиция: животное 1963 | S (6,28) | — | F | 12,68 | <0,0001 | |||||||
| S | ||||||||||||
| 905 1,00) | — | F | 58,45 | <0,0001 |
| Параметр . | edf . | Коэффициент . | ST . | Значение . | Вероятность . | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Предыдущее соотношение | — | 0,510 | т | 13,50 | <0,0001 | |||||||
| Животное 1932 | — | −1,025 | — | −1,025 | — | −1,025 <0,0001 | ||||||
| Животное 1963 | — | −0.788 | т | −11,32 | <0,0001 | |||||||
| Животное 1964 | — | −0,851 | т | −11,57 | −11,57 | — | F | 15,73 | <0,0001 | |||
| Положение: животное 1932 | S (4,88) | — | 905 01 | <0,0001 | ||||||||
| Позиция: животное 1963 | S (6,28) | — | F | 12,68 | <0,0001 | |||||||
| S | ||||||||||||
| 905 1,00) | — | F | 58,45 | <0,0001 |
Таблица 2. Результаты GAM
для отношений изотопов азота в вытяжке ( n = 163, скорректировано r 2 = 0 .983).
| Параметр . | edf . | Коэффициент . | ST . | Значение . | Вероятность . | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Предыдущее соотношение | — | 0,510 | т | 13,50 | <0,0001 | ||||||
| Животное 1932 | — | −1.041 | т | −12,38 | <0,0001 | ||||||
| Животное 1963 | — | −0,788 | т | −11,32 | −11,32 | −0,851 | т | −11,57 | <0,0001 | ||
| Положение: животное 1427 | S (3,26) | — | F | 15.73 | <0,0001 | ||||||
| Позиция: животное 1932 | S (4.88) | — | F | 8.01 | <0.0001 | ||||||
| Положение: | |||||||||||
| 6,28) | — | F | 12,68 | <0,0001 | |||||||
| Положение: животное 1964 | S (1,00) | — | F | 45 | <0,0001 |
| Параметр . | edf . | Коэффициент . | ST . | Значение . | Вероятность . | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Предыдущее соотношение | — | 0,510 | т | 13,50 | <0,0001 | ||||||
| Животное 1932 | — | −1.041 | т | −12,38 | <0,0001 | ||||||
| Животное 1963 | — | −0,788 | т | −11,32 | −11,32 | −0,851 | т | −11,57 | <0,0001 | ||
| Положение: животное 1427 | S (3,26) | — | F | 15.73 | <0,0001 | ||||||
| Позиция: животное 1932 | S (4.88) | — | F | 8.01 | <0.0001 | ||||||
| Положение: | |||||||||||
| 6,28) | — | F | 12,68 | <0,0001 | |||||||
| Положение: животное 1964 | S (1,00) | — | F | 45 | <0,0001 |
Таблица 3. Результаты GAM
для соотношений изотопов углерода в вытяжке [ n = 159, r 2 (прил.) = 0,98].
| Параметр . | edf . | Коэффициент . | ST . | Значение . | p -значение . | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Предыдущее отношение | — | −0.143 | т | −2,14 | 0,0343 | ||||||
| Отношение отставания 2 | — | 0,222 | т | 4,04 | <0,0001 | 4,04 | <0,0001 9025 - 905 - | <0,0001 905 - 905 - 905 1,090 | т | −8,23 | <0,0001 |
| Животное 1963 | — | −1,357 | т | −8.63 | <0,0001 | ||||||
| Животное 1964 | — | −0,824 | т | −7,51 | <0,0001 | ||||||
| Положение: животное 1427 | F | 15,93 | <0,0001 | ||||||||
| Положение: животное 1932 | S (7,07) | — | F | 13.42 | <0,0001 | ||||||
| Позиция: животное 1963 | S (7,79) | — | F | 11,54 | <0,0001 | ||||||
| Положение: животное 4.96) | — | F | 6,12 | <0,0001 |
| Параметр . | edf . | Коэффициент . | ST . | Значение . | p -значение . | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Предыдущее отношение | — | −0,143 | т | −2,14 | 0,0343 | ||||||
| Отношение отставания 2 | — | ||||||||||
| Животное 1932 | — | -1.090 | т | −8,23 | <0,0001 | ||||||
| Животное 1963 | — | -1,357 | т | -8,63 | -8,63 | −0,824 | т | −7,51 | <0,0001 | ||
| Положение: животное 1427 | S (7,88) | — | F | 15.93 | <0,0001 | ||||||
| Позиция: животное 1932 | S (7,07) | — | F | 13,42 | <0,0001 | ||||||
| S | |||||||||||
| 7,79) | — | F | 11,54 | <0,0001 | |||||||
| Положение: животное 1964 | S (4,96) | — | F 6. | <0,0001 |
Таблица 3. Результаты GAM
для соотношений изотопов углерода в вытяжке [ n = 159, r 2 (прил.) = 0,98].
| Параметр . | edf . | Коэффициент . | ST . | Значение . | p -значение . | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Предыдущее отношение | — | −0.143 | т | −2,14 | 0,0343 | ||||||
| Отношение отставания 2 | — | 0,222 | т | 4,04 | <0,0001 | 4,04 | <0,0001 9025 - 905 - | <0,0001 905 - 905 - 905 1,090 | т | −8,23 | <0,0001 |
| Животное 1963 | — | −1,357 | т | −8.63 | <0,0001 | ||||||
| Животное 1964 | — | −0,824 | т | −7,51 | <0,0001 | ||||||
| Положение: животное 1427 | F | 15,93 | <0,0001 | ||||||||
| Положение: животное 1932 | S (7,07) | — | F | 13.42 | <0,0001 | ||||||
| Позиция: животное 1963 | S (7,79) | — | F | 11,54 | <0,0001 | ||||||
| Положение: животное 4.96) | — | F | 6,12 | <0,0001 |
| Параметр . | edf . | Коэффициент . | ST . | Значение . | p -значение . | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Предыдущее отношение | — | −0,143 | т | −2,14 | 0,0343 | ||||||
| Отношение отставания 2 | — | ||||||||||
| Животное 1932 | — | -1.090 | т | −8,23 | <0,0001 | ||||||
| Животное 1963 | — | -1,357 | т | -8,63 | -8,63 | −0,824 | т | −7,51 | <0,0001 | ||
| Положение: животное 1427 | S (7,88) | — | F | 15.93 | <0,0001 | ||||||
| Позиция: животное 1932 | S (7,07) | — | F | 13,42 | <0,0001 | ||||||
| S | |||||||||||
| 7,79) | — | F | 11,54 | <0,0001 | |||||||
| Положение: животное 1964 | S (4,96) | — | F 6. | <0,0001 |
Согласованные изменения в наклонах соотношений между соотношением изотопов и положением вдоль клюва были менее очевидны из подобранных сглаживателей (не показаны), чем из необработанных данных, предположительно из-за того, что автокорреляция была удалена от бывшего.
Обсуждение
Профили δ 15 N подтвердили нашу гипотезу об обогащении 15 N в клюве по мере роста, что можно интерпретировать как свидетельство онтогенного изменения диеты.Этот вывод также подтверждает способность анализов стабильных изотопов отслеживать трофические изменения в диапазоне размеров от параларв до взрослых особей, как указывает Парри (2008). Наши результаты согласуются с результатами Хобсона и Черела (2006) и Руиса-Кули и др. . (2006) для других видов кальмаров. Тем не менее, в этих исследованиях использовались клювы разного размера, тогда как наш подход продемонстрировал, что одну и ту же информацию можно собрать с помощью одного клюва. Значения δ 15 N в верхней части рострального кончика соответствуют самым молодым фазам жизни гигантского кальмара, предполагая, что гигантский кальмар на ранних стадиях жизни питается на более низком трофическом уровне, чем взрослые и взрослые особи.
Как и ожидалось, поскольку δ 15 N является хорошим индикатором трофического положения (DeNiro, Epstein, 1981), мы наблюдали различия в значениях δ 15 N в пределах одного и того же верхнего клюва, увеличиваясь от рострума к задней части. край. Этот результат эквивалентен результатам Cherel and Hobson (2005) при сравнении значений δ 15 N в разных частях одного и того же клюва.
Обогащение δ 15 N, наблюдаемое между верхушкой рострального конца и задним краем капюшона, варьировалось в пределах 5.2 и 7,4 ‰, чуть менее чем двукратная разница между трофическим положением малого и большого A. dux . Cherel и др. . (2009b) обнаружили, что клювы взрослых гигантских кальмаров были более обогащены δ 15 N, чем клювы молоди на 1,7 ‰, то есть примерно на половину трофического уровня. Однако, поскольку клюв объединяет материал, отложившийся на всех этапах жизни животного, сравнение целых клювов будет иметь тенденцию недооценивать трофические различия. Онтогенетическое различие двух трофических уровней находится в пределах, обнаруженных у особей O.bartramii и S. oualaniensis (Parry, 2008) и T. filippovae (Cherel et al ., 2009a).
Значения δ 15 N, обнаруженные вдоль заднего края верхнего клюва созревающего гигантского кальмара (9,9–12,3 ‰) из Бискайского залива (Приложение), могут свидетельствовать о том, что этот вид будет самым большим высшим хищником в мезопелагическая экосистема района. Однако следует проявлять осторожность при интерпретации данных таким образом, поскольку значения δ 15 N из нижних клювов взрослого человека A.dux в том же районе показал несколько более низкие значения (∼8,5–10,8 ‰), а главным хищником кальмаров в этой мезопелагической экосистеме является Taningia danae (Cherel et al ., 2009b).
Проанализированные нами верхние клювы показали, что значения δ 13 C увеличились с -17,8 до -15,8 ‰ от вершины рострума до заднего края капюшона у трех испанских особей и с -16,8 до -14,3 в дюймах. намибийский. Аналогичная тенденция к увеличению δ 13 C была обнаружена Cherel et al .(2009a) в нижнем клюве T. filippovae , причем самые недавно сформированные области (крыло и боковая стенка) более обогащены δ 13 C, чем более старые области. Значения δ 13 C, полученные Cherel et al . (2009b) в нижних крыльях клюва взрослой особи A. dux , на которую охотятся кашалоты в Бискайском заливе (-16,6 ± 0,4 ‰), были сопоставимы с таковыми из того же района (-16,2 ± 0,4 ‰), но были отличается от намибийского образца (-14,9 ± 0.6 ‰).
Поскольку δ 13 C обычно обогащается примерно на 1 ‰ на трофический уровень, эти результаты согласуются с результатами для δ 15 N, предполагая, что A. dux повышается примерно на два трофических уровня за время своего существования; Наблюдаемые различия в жизни животных также могут быть результатом перемещения между разными районами кормодобывания с разными первичными источниками углерода (ДеНиро и Эпштейн, 1978; Хобсон и др. ., 1994; Черел и др. ., 2000).Колебания профилей δ 13 C около верхушки рострального слоя (рис. 3) могут быть связаны с большей внутренней изменчивостью изотопного состава углерода относительно небольшой добычи (Cabana and Rasmussen, 1996). С другой стороны, различия могут быть связаны с миграционным поведением кальмаров на раннем этапе их жизни.
Относительная стабильность в профилях δ 13 C после ∼20 мм вдоль клюва (рис. 3) может быть связана с более постоянным и определенным пищевым ресурсом в более позднем возрасте.Согласованность значений изотопов углерода также может означать, что взрослые гигантские кальмары живут в одном районе, питаясь добычей с аналогичным изотопным составом углерода. Сидячий образ жизни соответствовал бы плохим способностям плавания Architeuthis (Roper and Boss, 1982; Guerra et al., ., 2006). Для поддержания высокой скорости роста, характерной для этих животных (Grist and Jackson, 2007), при относительно низкой эффективности передвижения и относительно низком метаболизме (Seibel et al ., 2000), Architeuthis должен жить в областях с относительно высокой производительностью. Места отлова Architeuthis были относительно близки к подводным каньонам и водным массам, характеризующимся высокой продуктивностью (Guerra et al ., 2005).
Различия в биохимическом составе клювов могут затруднить интерпретацию различий в изотопных значениях. Различное соотношение хитина к белку было обнаружено в незатемненных, затемненных и затемненных частях клюва кальмаров, причем в непрозрачных частях хитина было больше, чем в затемненных (Miserez et al ., 2008). Поскольку хитин обеднен δ 15 N по сравнению с диетой и имеет более высокое отношение C / N, чем белок (6,9 против 3–4; Schimmelmann and DeNiro, 1986; Webb et al. ., 1998), тем выше чем выше содержание хитина, тем ниже значение δ 15 N и выше отношение C / N. В нашем исследовании мы обнаружили, что значения отношения масс C / N были ниже (1,99–1,42; Приложение), чем у других кальмаров (например, 3,45–3,60 у T. filippovae ; Cherel et al ., 2009a). . Это может указывать на то, что клюв Architeuthis темнее, чем у T.filippovae . С другой стороны, отношение C / N уменьшается от рострума к краю клюва верхнего капюшона, что означает, что эта часть богаче хитином, чем рострум. Это подчеркивало тот факт, что край капюшона отслеживает недавнее кормление.
Совпадает с выводами Такаи и др. . (2000), различия в значениях δ 13 C между испанскими и намибийскими образцами могут просто указывать на то, что первые населяли географические области, отличные от последних.Достоверное прямое сравнение значений δ 15 N особей из разных районов требует вычитания базовых уровней из значений δ 15 N хищников, поскольку исходные значения варьируются в зависимости от экосистем.
Благодарности
Мы благодарим Ива Шереля и двух анонимных рецензентов за их комментарии и предложения, которые существенно улучшили статью. Исследование финансировалось за счет премии (DE-FC09-96-SR18546) Министерства энергетики США Экологической лаборатории Саванна-Ривер Университета Джорджии.GJP финансировался проектом ANIMATE (MEXC-CT-2006-042337).
Список литературы
,,,.
Реконструкция трофических путей между планктоном и северной иберийской сардиной ( Sardina pilchardus ) с использованием стабильных изотопов
,
Scientia Marina
,
2004
, vol.
68
(стр.
165
—
178
),.
Головоногие моллюски: экология и рыболовство
,
2005
Оксфорд, Великобритания
Blackwell Science
стр.
452
,.
Сравнение водных пищевых цепей с использованием изотопов азота
,
Proceedings of the National Academy of Sciences of the USA
,
1996
, vol.
93
(стр.
10844
—
10847
),,,,.
Тканевые, онтогенные и связанные с полом различия в значениях δ 13 C и δ 15 N океанических кальмаров Todarodes filippovae (Cephalopoda: Ommastrephidae)
,
Marine Biology
,
2009
, vol.
156
(стр.
699
—
708
),.
Стабильные изотопы, клювы и хищники: новый инструмент для изучения трофической экологии головоногих моллюсков, включая гигантских и колоссальных кальмаров
,
Труды Лондонского королевского общества, серия B
,
2005
, vol.
272
(стр.
1601
—
1607
),.
Географические различия в сигнатурах стабильных изотопов углерода морских хищников: инструмент для исследования ареалов их кормления в Южном океане
,
Marine Ecology Progress Series
,
2007
, vol.
329
(стр.
281
—
287
),,.
Использование стабильного изотопного анализа перьев для определения происхождения морских птиц после линьки и размножения
,
Oecologia
,
2000
, vol.
122
(стр.
155
—
162
),,,.
Стабильные изотопы документируют трофическую структуру комплекса глубоководных головоногих моллюсков, включая гигантского осьминога и гигантского кальмара
,
Biological Letters
,
2009
, vol.
5
(стр.
364
—
367
).
Справочник по идентификации клювов головоногих
,
1980
Oxford
Clarendon Press
стр.
273
,,,.
Генетический скрининг содержимого кишечника гигантского кальмара ( Architeuthis sp.)
,
Journal of Heredity
,
2005
, vol.
96
(стр.
417
—
423
),.
Влияние диеты на распределение изотопов углерода у животных
,
Geochimica et Cosmochimica Acta
,
1978
, vol.
42
(стр.
495
—
506
),.
Влияние диеты на распределение изотопов азота у животных
,
Geochimica et Cosmochimica Acta
,
1981
, vol.
45
(стр.
341
—
351
),,,,.
Возрастные сдвиги в составе рациона южных морских слонов расширяют общую нишу кормодобывания
,
Морская биология
,
2007
, vol.
150
(стр.
1441
—
1452
),,,.
Недавние находки гигантского кальмара Architeuthis в северных водах Испании
,
Журнал Морской биологической ассоциации Великобритании
,
2002
, vol.
82
(стр.
859
—
861
),.
Сколько времени нужно, чтобы стать гигантским кальмаром?
,
Обзоры по биологии рыб и рыболовству
,
2007
, vol.
17
(стр.
385
—
399
),,,.
Обзор записей гигантских кальмаров в северо-восточной части Атлантического океана с примечанием к двум первым записям самцов. Architeuthis sp.от Пиренейского полуострова
,
Журнал Морской биологической ассоциации Великобритании
,
2004
, vol.
84
(стр.
427
—
431
),,,,.
Enigmas de la ciencia: el calamar gigante
,
2006
Fundación Caja Rural de Asturias (Овьедо, Испания) и Instituto de Investigaciones Marinas, CSIC, Виго, Испания
стр.
313
,,,.
Architeuthis записей и взаимосвязей с океанскими процессами и топографией дна
,
2005
ASLO Summer 2005 Meeting, Book of Abstracts: 65.
,,.
Возраст определен по ежедневному отложению концентрических колец на клювах обыкновенного осьминога ( Octopus vulgaris )
,
Fishery Bulletin US
,
2001
, vol.
99
(стр.
679
—
684
),,.
Определение возраста и роста мелкотелого осьминога из Новой Зеландии
,
2009
Симпозиум Международного консультативного совета головоногих моллюсков, Виго, 3–11 сентября 2009 г., Книга рефератов: 76.
.
Анализ стабильных изотопов мраморных кайрелей: свидетельство питания пресной водой и определение трофического уровня
,
The Condor
,
1990
, vol.
92
(стр.
897
—
903
),.
Изотопная реконструкция морских пищевых сетей с использованием клювов головоногих моллюсков: новый взгляд на выращивание в неволе Sepia officinalis
,
Canadian Journal of Zoology
,
2006
, vol.
84
(стр.
766
—
770
),,.
Использование стабильных изотопов для определения трофических отношений морских птиц
,
Journal of Animal Ecology
,
1994
, vol.
63
(стр.
786
—
798
),.
Сравнительное исследование белкового состава хитин-белковых комплексов клюва, пера, присоски, радулы и кутикулы пищевода головоногих
,
Сравнительная биохимия и физиология
,
1981
, vol.
68B
(стр.
535
—
546
),,,,,, и др.
Применение новых инструментов к трофической динамике и экологии головоногих: перспективы семинара по головоногим в Южном океане, 2–3 февраля 2006 г.
,
Обзоры по биологии и рыболовству рыб
,
2007
, vol.
17
(стр.
79
—
99
),,,.
Изменение трофического сдвига для соотношений стабильных изотопов углерода, азота и серы
,
Oikos
,
2003
, т.
102
(стр.
378
—
390
),,,,.
δ 13 C и δ 15 N профилирование зубов кашалота выявляет онтогенетические движения и трофическую экологию
,
Oecologia
,
2007
, vol.
151
(стр.
605
—
615
),,,,.
Переход от жестких материалов к податливым в клювах кальмаров
,
Science
,
2008
, vol.
319
(стр.
1816
—
1819
). .
Диеты головоногих моллюсков
,
Жизненные циклы головоногих моллюсков. 2. Сравнительные обзоры
,
1987
Лондон
Academic Press
(стр.
201
—
219
).
Трофическая изменчивость в зависимости от длины у двух кальмаров оммастрофид, Ommastrephes bartramii и Sthenoteuthis oualaniensis
,
Marine Biology
,
2008
, vol.
153
(стр.
249
—
256
),
Гарсия-Сантамария
M. T.
,,,.
Сравнение возраста чтения сагиттальных сечений и боковых стенок клювов осьминога ( Octopus vulgaris Cuvier, 1797)
,
2009
Симпозиум Международного консультативного совета головоногих моллюсков, Виго, 3–11 сентября 2009 г., Книга рефератов: 55.
,,.
Планктон 13 C: 12 Соотношение C меняется с широтой: разница между северным и южным океанами
,
Deep Sea Research A
,
1982
, vol.
29
(стр.
1035
—
1039
),
Эрнандес-Гонсалес
C. L.
.
Линии роста в микроструктуре клюва осьминога Octopus vulgaris Cuvier, 1797. Cephalopod Biodiversity, Ecology and Evolution
,
South African Journal of Marine Science
,
1998
, vol.
20
(стр.
135
—
142
),.
Роль потребителей
,
Философские труды Лондонского королевского общества, серия B
,
1996
, vol.
351
(стр.
1003
—
1022
),.
Гигантский кальмар
,
Scientific American
,
1982
, т.
246
(стр.
96
—
105
),,,,.
Трофические отношения между кашалотами и гигантскими кальмарами с использованием стабильных изотопов C и N
,
Marine Ecology Progress Series
,
2004
, vol.
277
(стр.
275
—
283
),,,.
Стабильные изотопы в клювах гигантского кальмара ( Dosidicus gigas ) для оценки его трофического положения: сравнение содержимого желудка и стабильных изотопов
,
Журнал Морской биологической ассоциации Великобритании
,
2006
, vol.
86
(стр.
437
—
445
),.
Исследования стабильных изотопов хитина. 2. Отношения 13 C / 12 C и 15 N / 14 N в хитине членистоногих
,
Contributions to Marine Science
,
1986
, vol.
29
(стр.
113
—
130
),,.
Ограничение света во взаимодействиях хищник-жертва: последствия для метаболизма и передвижения глубоководных головоногих моллюсков
,
Биологический бюллетень
,
2000
, vol.
198
(стр.
284
—
298
),,,.
Различение популяций пресноводных и морских морских котиков ( Phoca vitulina ) с использованием соотношений стабильных изотопов и профилей жирных кислот
,
Canadian Journal of Fisheries and Aquatic Sciences
,
1996
, vol.
53
(стр.
272
—
279
),,,,.
Экспериментальное исследование влияния диеты на профили жирных кислот и стабильных изотопов кальмаров Lolliguncula brevis
,
Journal of Experimental Marine Biology and Ecology
,
2006
, vol.
333
(стр.
97
—
114
),,,,,.
Географические вариации в соотношении стабильных изотопов углерода и азота у кальмаров
,
Журнал Морской биологической ассоциации Великобритании
,
2000
, vol.
80
(стр.
675
—
684
),.
Источники различий в рационе потребителей δ 15 Обогащение N: метаанализ
,
Oceanologia
,
2003
, vol.
136
(стр.
32
—
37
),,.
Качество рациона влияет на δ 13 C и δ 15 N саранчи и их биохимические компоненты
,
Journal of Experimental Biology
,
1998
, vol.
201
(стр.
2903
—
2911
),,.
Анализ экологических данных. Статистика для биологии и здоровья. Серия
,
2007
Берлин
Springer
стр.
685
Приложение
δ 15 N и δ 13 Значения C в капюшоне верхних клювов Architeuthis из Бискайского залива (Астурия, северная Испания, номера экземпляров 1932, 1964 и 1963) и воды Намибии (1427) .D — расстояние от кончика рострального клюва до заднего края верхнего клюва, где отбиралась каждая проба; Среднее значение отношения C / N и стандартное отклонение.
| . | 1932 . | 1964 . | 1963 . | 1427 . | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| D (мм) . | δ 15 с.ш. . | δ 13 С . | δ 15 с.ш. . | δ 13 С . | δ 15 с.ш. . | δ 13 С . | δ 15 с.ш. . | δ 13 С . | ||||||||||||
| 1,43 | 5,48 | −17,21 | 5,27 | −17,16 | 5,61 | −17,81 | 5,98 | −16.94 | ||||||||||||
| 2,86 | 6,12 | −16,96 | 6,38 | −16,03 | 6,22 | −17,43 | 7,35 | −15,81 | 7,45 | −15,81 | −15,81 | −16,86 | 7,24 | −16,76 | 7,84 | −15,46 | ||||
| 5,72 | 6,84 | −16,54 | 8,11 | 905,02 | −16,76 | 8,39 | −15,49 | |||||||||||||
| 7,15 | 7,61 | −16,52 | 8,46 | −16,20 | 8,33 | −16,20 | 8,33 | −16,20 | 8,33 | −16,20 | 8,33 | -16,20 | 8,19 | −16,82 | 8,57 | −16,14 | 8,82 | −16,79 | 9,17 | −15,23 |
| 10,00 | 825 −1 | 8,49 | −16,15 | 8,53 | −16,64 | 9,53 | −15,36 | |||||||||||||
| 11,43 | 8,62 | −16,56 | 8,62 | −16,56 | 8,62 | −16,56 | −15,35 | |||||||||||||
| 12,86 | 8,65 | −16,59 | 8,86 | −16,08 | 8,95 | −17,01 | 9,825 − | |||||||||||||
| 14,29 | 8,28 | -17,02 | 8,91 | -15,95 | 9,21 | -17,07 | 9,96 | -15,36 | 9,96 | -15,36 | −15,93 | 8,93 | −16,58 | 10,19 | −15,23 | |||||
| 17,15 | 8,88 | −16,60 | 8,94 | −15.88 | 9,41 | −16,70 | 10,28 | −15,20 | ||||||||||||
| 18,58 | 9,03 | −16,55 | 9,08 | −16,02 | 9,08 | −16,02 | 9,425 9,425 9,425 | |||||||||||||
| 20,01 | 9,09 | -16,52 | 9,23 | -15,86 | 9,44 | -16,71 | 10,39 | -15,07 | ||||||||||||
| 21.44 | 9,17 | -16,62 | 9,25 | -15,88 | 9,23 | -16,62 | 10,49 | -15,01 | ||||||||||||
| 22,87 | 22,87 | −16,54 | 10,57 | −15,01 | ||||||||||||||||
| 24,30 | 9,43 | −16,31 | 9,38 | −15,83 | 9,57 −1 | 10,61 | −14,89 | |||||||||||||
| 25,73 | 9,38 | −16,21 | 9,50 | −15,86 | 9,54 | −16,53 | 9,54 | −16,53 | −16,53 | −16,24 | 9,45 | −15,80 | 9,74 | −16,39 | 10,92 | −15,02 | ||||
| 28,59 | 9,29 | −16.05 | 9,55 | −15,82 | 9,47 | −16,52 | 11,03 | −15,04 | ||||||||||||
| 30,02 | 9,27 | −16,05 | 9,27 | −16,05 | 9,65 11,25 | −14,90 | ||||||||||||||
| 31,45 | 9,36 | −15,99 | 9,69 | −15,79 | 9,97 | −16,42 | 11,33 −14526 | |||||||||||||
| 32,88 | 9,52 | −16,03 | 9,75 | −15,72 | 10,16 | −16,32 | 11,40 | −14,89 | −14,89 | −15,81 | 10,15 | −16,29 | 11,42 | −14,90 | ||||||
| 35,74 | 9,67 | −16,05 | 9,87 | −15.71 | 10,31 | −16,30 | 11,51 | −14,79 | ||||||||||||
| 37,17 | 9,62 | −16,05 | 10,01 | −15,72 26 | 1125,65 | −15,72 26 | 10,25 | |||||||||||||
| 38,60 | 9,66 | -16,05 | 10,06 | -15,70 | 10,39 | -16,29 | 11,81 | -14,58 | ||||||||||||
| 40.03 | 9,78 | −15,95 | 10,24 | −15,72 | 10,42 | −16,27 | 12,10 | −14,52 | ||||||||||||
| 41,46 | −15,72 | 41,46 | −5 10,51 | -16,20 | 12,28 | -14,37 | ||||||||||||||
| 42,89 | 9,96 | -15,96 | 10,21 | -15,64 | 10,525 -16 | 12,54 | -14,38 | |||||||||||||
| 44,32 | 10,17 | -15,94 | 10,23 | -15,66 | 10,57 | -16,29 ,38 | 10,57 | -16,29 ,38 45265 | −15,73 | 10,34 | −15,64 | 10,53 | −16,26 | 12,73 | −14,26 | |||||
| 47,18 | 10,28 | −15.78 | 10,44 | −15,50 | 10,62 | −16,32 | 12,59 | −14,16 | ||||||||||||
| 48,61 | 10,38 | −15,59 | −1 | −15,59 | −1 | −14,07 | ||||||||||||||
| 50,04 | 10,37 | −15,57 | 10,58 | −15,55 | 10,55 | −16,20 | 12,97 −13526 | 12,97 −13526 | ||||||||||||
| 51,47 | 10,69 | −15,47 | 10,63 | −15,72 | 10,68 | −16,19 | 13,45 | −14,05 | −14,05 | −14,05 | −15,75 | 10,56 | −16,29 | 13,54 | −13,94 | |||||
| 54,33 | 9,98 | −15,56 | 10,78 | −15.63 | 10,65 | −16,20 | 13,45 | −14,34 | ||||||||||||
| 55,76 | 10,17 | −15,58 | 10,84 | −15,58 | 10,84 | −15,95 | −15,95 | 9,81 | −15,73 | 10,68 | −15,76 | 10,61 | −16,26 | |||||||
| 58,62 | 9.96 | -15,83 | 10,76 | -16,25 | ||||||||||||||||
| 60,05 | 9026 | 1025 905 905 905 905 9025 9025 | 11,14 | −16,11 | ||||||||||||||||
| 62,91 | 11.34 | −15,97 | ||||||||||||||||||
| 64,34 | 11,41 | 9026 9026 905 905 905 905 905 905 905 905 9025 905 905 | −16,01 | |||||||||||||||||
| 67,20 | 12,02 | −16.22 | ||||||||||||||||||
| 68,63 | 12,32 | -16,43 | ||||||||||||||||||
| 1,42 ± 0,35 | ||||||||||||||||||||
| . | 1932 . | 1964 . | 1963 . | 1427 . | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| D (мм) . | δ 15 с.ш. . | δ 13 С . | δ 15 с.ш. . | δ 13 С . | δ 15 с.ш. . | δ 13 С . | δ 15 с.ш. . | δ 13 С . | ||||||||
| 1,43 | 5,48 | −17,21 | 5,27 | −17,16 | 5,61 | −17,81 | 5,98 | −17,81 | 5,98 | −16,9425 6,35 | −16,03 | 6,22 | −17,43 | 7,35 | −15,81 | |
| 4,29 | 7,49 | −16,51 | 7,87 | −16.86 | 7,24 | −16,76 | 7,84 | −15,46 | ||||||||
| 5,72 | 6,84 | −16,54 | 8,11 | −16,02 | 8,11 | −16,02 −5 | ||||||||||
| 7,15 | 7,61 | −16,52 | 8,46 | −16,20 | 8,33 | −16,45 | 8,77 | −15,40 | ||||||||
| 8,5258 90.19 | −16,82 | 8,57 | −16,14 | 8,82 | −16,79 | 9,17 | −15,23 | |||||||||
| 10,00 | 8,01 | −165 ,526 −16,64 | 9,53 | −15,36 | ||||||||||||
| 11,43 | 8,62 | −16,56 | 8,78 | −15,99 | 8,84 | −16,725 9 905.63 | −15,35 | |||||||||
| 12,86 | 8,65 | −16,59 | 8,86 | −16,08 | 8,95 | −17,01 | 9,82 | 8,25 | 8,91 | −15,95 | 9,21 | −17,07 | 9,96 | −15,36 | ||
| 15,72 | 8,95 | −16,56 | 8.91 | −15,93 | 8,93 | −16,58 | 10,19 | −15,23 | ||||||||
| 17,15 | 8,88 | −16,60 | 8,94 | −16,60 | 8,94 | 8259 | ||||||||||
| 18,58 | 9,03 | −16,55 | 9,08 | −16,02 | 9,44 | −16,55 | 10,35 | −15,14 | 9,09 | -16,52 | 9,23 | -15,86 | 9,44 | -16,71 | 10,39 | -15,07 | 21,44,17 -5 9,23 | −16,62 | 10,49 | −15,01 |
| 22,87 | 9,31 | −16,28 | 9,33 | −15,84 | 9,62 −16526 | 9,62 −16526 | 10,57 | −15,01 | ||||||||
| 24,30 | 9,43 | −16,31 | 9,38 | −15,83 | 9,57 | −16,46 −1 | −16,46 −5 | −16,21 | 9,50 | −15,86 | 9,54 | −16,53 | 10,79 | −14,95 | ||
| 27,16 | 9,39 | −16.24 | 9,45 | −15,80 | 9,74 | −16,39 | 10,92 | −15,02 | ||||||||
| 28,59 | 9,29 | −16,05 −1 | −16,05 −7 11,03 | −15,04 | ||||||||||||
| 30,02 | 9,27 | −16,05 | 9,67 | −15,80 | 9,90 | −16,40 | 11,25 | −14.90 | ||||||||
| 31,45 | 9,36 | −15,99 | 9,69 | −15,79 | 9,97 | −16,42 | 11,33 | −16,42 | 11,33 | −14,94 25 905 | −15,72 | 10,16 | −16,32 | 11,40 | −14,89 | |
| 34,31 | 9,51 | −15,98 | 10,01 | −15.81 | 10,15 | −16,29 | 11,42 | −14,90 | ||||||||
| 35,74 | 9,67 | −16,05 | 9,87 | −15,71 | −15,71 | 10 | ||||||||||
| 37,17 | 9,62 | -16,05 | 10,01 | -15,72 | 10,27 | -16,12 | 11,63 | -14,79 | ||||||||
| 38.60 | 9,66 | −16,05 | 10,06 | −15,70 | 10,39 | −16,29 | 11,81 | −14,58 | ||||||||
| 40,03 | 9,78 10,42 | −16,27 | 12,10 | −14,52 | ||||||||||||
| 41,46 | 9,95 | −15,92 | 10,20 | −15,72 | 10,525 −1 | 12,28 | −14,37 | |||||||||
| 42,89 | 9,96 | −15,96 | 10,21 | −15,64 | 10,54 | −16,27 12,526 | −16,27 12,526 | -15,94 | 10,23 | -15,66 | 10,57 | -16,29 | 12,50 | -14,37 | ||
| 45,75 | 10,26 | -15.73 | 10,34 | −15,64 | 10,53 | −16,26 | 12,73 | −14,26 | ||||||||
| 47,18 | 10,28 | −15,7255 | −15,7252 | −15,725 10254 | −15,725 10254 12,59 | −14,16 | ||||||||||
| 48,61 | 10,38 | −15,59 | 10,52 | −15,66 | 10,61 | −16,31 | 12,625 −1 | |||||||||
| 50,04 | 10,37 | −15,57 | 10,58 | −15,55 | 10,55 | −16,20 | 12,97 | −13,88 | −13,88 | −15,72 | 10,68 | −16,19 | 13,45 | −14,05 | ||
| 52,90 | 10,59 | −15,41 | 10,69 | −15.75 | 10,56 | −16,29 | 13,54 | −13,94 | ||||||||
| 54,33 | 9,98 | −15,56 | 10,78 | −15,63 −15,63 | −15,63 | |||||||||||
| 55,76 | 10,17 | -15,58 | 10,84 | -15,95 | 10,64 | -16,35 | ||||||||||
| 57,19 | −15,73 | 10,68 | −15,76 | 10,61 | −16,26 | |||||||||||
| 58,62 | 9,96 | |||||||||||||||
| 60,05 | 10,99 | −16,06 | ||||||||||||||
| 61.48 | 11,14 | −16,11 | ||||||||||||||
| 62,91 | 11,41 | −16,03 | ||||||||||||||
| 65,77 | 11.57 | −16,01 | ||||||||||||||
| 67.20 | 12.02 | −16.22 | −16,43 | |||||||||||||
| C / N | 1,99 ± 1,25 | 1,72 ± 0,33 | 1,71 ± 0,34 | 1.42 ± 0,35 | ||||||||||||
© 2010 Международный совет по исследованию моря. Опубликовано Oxford Journals. Все права защищены. Для получения разрешений обращайтесь по электронной почте: [email protected]
.
Первое сообщение о мертвом гигантском кальмаре (Architeuthis dux) с работающего сейсмического судна | Записи о морском биоразнообразии
Виды гигантских кальмаров, будь то в море или выброшенные на берег, редки, а распространение этого вида остается плохо изученным (Roper and Shea 2013).Большинство записей Architeuthis происходит из Северной Атлантики, несколько сообщений — из южной части Бразилии (Arfelli et al. 1991; Martins and Perez, 2009). Наши данные дополняют ограниченную информацию о распространении этого вида в южной части Атлантического океана.
Поскольку у нас не было возможности исследовать животное, мы понимаем, что его смерть не может быть окончательно связана с сейсмической активностью. Тем не менее мы считаем, что его данные помогают выявить вызывающую тревогу нехватку информации о воздействии звукового загрязнения морской среды на беспозвоночных.
До недавнего времени опасения по поводу антропогенного шума в морской экосистеме были связаны исключительно с его воздействием на звук как средство коммуникации, игнорируя влияние звука как волны давления, которая может причинить физический вред определенным видам (André et al. 2011 г.). Поэтому законодательство было разработано для защиты животных, которые, как и люди, полагаются на звук для общения, и не смогли защитить животных, которые воспринимают звук иначе. Хотя нет никаких определенных доказательств связи гибели этого экземпляра с сейсмической активностью, его появление в районе активных исследований наводит на мысль о возможной связи.Наблюдения за гигантскими кальмарами в море редки, и существует правдоподобный механизм, позволяющий ожидать, что использование пневматического оружия может вызвать физические травмы. Мы считаем, что принцип предосторожности следует применять в подобных ситуациях, когда виды могут подвергнуться опасности в результате деятельности человека, до тех пор, пока не будет достигнуто лучшее научное понимание.
С юридической точки зрения государства несут общие обязательства по международному праву по защите и сохранению морской среды и устранению конкретных источников загрязнения морской среды (Dotinga and Elferink 2000), включая акустическое загрязнение.Общая обязанность, закрепленная в статье 192 Конвенции Организации Объединенных Наций по морскому праву, которую ратифицировала Бразилия, включает обязанность защищать морскую среду от воздействия подводного шума, которая охватывается определением, поскольку включает введение энергии в морскую среду, что может привести к пагубным последствиям (Firestone and Jarvis 2007).
Бразилия уже предприняла усилия по защите критических местообитаний морских млекопитающих от акустических нарушений.От Бразилии требуется принять превентивные меры, основанные на предупредительном и упреждающем подходах. Это обязательство распространяется как на любую деятельность, осуществляемую в исключительной экономической зоне Бразилии, так и на любые суда, плавающие в открытом море под бразильским флагом. Создание правил, которые способствуют сбору дополнительных морских научных данных в этой области и которые учитывают потенциальное воздействие гидролокатора на беспозвоночных, таких как глубоководные кальмары, было бы способом выполнить это обязательство.
В свете этих юридических обязательств и неопределенностей, связанных с воздействием на беспозвоночных, мы считаем, что регулирующим органам срочно необходимо провести всесторонний анализ воздействия сейсмических исследований на различные таксоны, а также на морских млекопитающих. Если нефтегазовые компании действительно привержены защите окружающей среды, они также должны внести свой вклад в предоставление ресурсов и содействие таким усилиям.
Чрезвычайное количество гигантских кальмаров Architeuthis dux, обнаруженных в японских прибрежных водах Японского моря с января 2014 г. по март 2015 г.
Японское море — это окраинное море протяженностью 978 000 км. 2 , расположенное на западной периферии. северная часть Тихого океана.Он граничит с Евразийским континентом на западе и Японским архипелагом на востоке и связан с Охотским морем через узкий Соевый пролив на севере, с северной частью Тихого океана через пролив Цугару на востоке и с Восточным Китаем. Море через узкий Цусимский пролив на юг. Средняя глубина составляет около 1667 м, а самая глубокая точка — 3742 м. Теплое Цусимское течение впадает в Японское море через Цусимский пролив с юга, проходит вдоль побережья Японии на север и вытекает через пролив Цугару в северную часть Тихого океана и через пролив Соя в Охотское море.Из-за узкой (ширина около 200 км) и неглубокой (глубина 90–130 м) географии Цусимского пролива ограниченная циркуляция воды происходит в верхних слоях, глубина которых ниже ок. Глубина 300 м. Еще глубже так называемая «Собственная вода Японского моря», которая характеризуется чрезвычайно холодной (0–1 ° C) и высокой соленостью (34,1 PSU), распространяется по всему Японскому морю (http: // www. jma-net.go.jp/jsmarine/japansea.html).
По оценкам, гигантские кальмары обитают в мезопелагических водах открытых океанов умеренного пояса (Clarke 1966; Roper and Boss 1982).Кубодера и Мори (2005) обнаружили, что гигантский кальмар у островов Огасавара в северной части Тихого океана появился на глубине 900 м и проплыл до 600 м, чтобы спастись от приманки, на которую было зацеплено его щупальце. Они сообщили, что температура воды на глубине 900 и 600 метров составляла около 4 и 6 ° C соответственно. Эти данные свидетельствуют о том, что гигантские кальмары в Японском море могут быть не постоянными жителями, а скорее мигрантами с юга, проходящими через воды придонного слоя Цусимского пролива, когда температура воды там опускается ниже ок.6 ° C с середины зимы до начала весны. После того, как они попали в Японское море, температура воды в глубоком слое становится слишком низкой, и они, вероятно, перемещаются через толщу воды в более подходящую температурную зону между теплым поверхностным слоем и холодной водой Японского моря в течение лета до осени.
Okiyama (1993) предложил возможную причину того, почему гигантских кальмаров иногда находили мертвыми на пляжах или ловили в стационарных сетях, установленных в прибрежных водах Японского моря в период с зимы до начала весны.В начале зимы поверхностные воды охлаждаются с северных периферий вдоль континентального побережья, и холодные поверхностные воды при температуре 0–1 ° C опускаются в более глубокий пласт, который является источником собственно воды Японского моря. Проседание холодной воды начинается с севера и постепенно распространяется на юг по мере продвижения зимы. Это движение холодной воды может снизить подходящую температуру среды обитания гигантского кальмара как по горизонтали, так и по вертикали. Они вынуждены двигаться на юг и на более мелкие глубины.Некоторые люди будут ослаблены воздействием чрезвычайно холодной воды, а затем перенесены вихрями Цусимского течения и сильными северо-западными сезонными ветрами, что приведет к их приближению к прибрежным водам Японии и объяснит, что они оказались в ловушке в стационарных сетях и / или оказались на мели. на пляжах японского побережья.
Хотя это одно из возможных объяснений появления гигантских кальмаров в японских прибрежных водах Японского моря, необычные массовые находки гигантских кальмаров в 2014 и 2015 годах, вероятно, связаны с другим механизмом.Судя по 10-дневным и месячным аномалиям средней температуры воды у поверхности и на глубине 100 м в зимние сезоны 2014 и 2015 годов, температура воды на этих глубинах была явно ниже в оба года, чем обычно, особенно в январе 2014 и феврале 2015 года. Заметные крупномасштабные холодные водные массы, которые образовались в среднем слое центральной части Японского моря в 2014 и 2015 годах, возможно, сыграли важную роль в переносе гигантского кальмара на поверхность и к побережью Японии.Кроме того, хотя в период 2008–2013 гг. Было немного записей о гигантских кальмарах (Кубодера, личное наблюдение), вполне возможно, что некоторое количество гигантских кальмаров проникло в Японское море в эти годы и выжило там до 2014 г.
Один из них. Важными биологическими находками гигантских кальмаров, сделанными на основании недавних массовых открытий, является бимодальное частотное распределение размеров (80–160 см DML с модой 110 см и 160–190 см DML с модой 170 см DML) в зимний сезон. В меньшей группе соотношение полов было почти равномерным, а большая группа полностью состояла из женщин.Wada et al. (2015) недавно сообщили, что 14 июня 2013 г. два молодых гигантских кальмара размером 33 см DML были пойманы кошельковой неводной сетью в юго-западной части Японского моря. В январе месячный темп роста составит ок. 11 см DML. Это значение представляет очень быстрый рост и склонность к S-образной кривой роста с более низкими темпами роста на ранних и поздних стадиях жизни. Общий размерный состав гигантских кальмаров в Японском море предполагает продолжительность жизни самцов 2 года и самок 3 года.
Ропер и Ши (2013) проанализировали современные знания о таксономии и систематике, распределении, размере популяции, использовании среды обитания, возрасте и росте, хищничестве и кормлении, воспроизводстве и жизненных циклах, а также функциональной морфологии гигантских кальмаров и обнаружили большие пробелы. Настоящие массовые находки 57 особей за относительно короткий период времени, два зимних сезона в период с 2014 по 2015 год и ограниченный район японского побережья Японского моря значительно увеличили доступную информацию, необходимую для понимания естественной истории гигантских кальмаров. .
Roper et al. (2015) исследовали записи гигантских кальмаров, обнаруженных в западной части Северной Атлантики между Ньюфаундлендом и Мексиканским заливом, и предоставили подробную информацию о 28 особях, обнаруженных в период 1952–2011 годов. Они признали общую тенденцию к увеличению числа наблюдений в 1990-х и 2000-х годах, что отражает возросшую научную осведомленность и растущий интерес населения к гигантским кальмарам. В Японии в 2013 году у нас было два крупных мероприятия, посвященных гигантскому кальмару. Одно из них — телевизионная программа, транслировавшаяся в январе национальной общественной вещательной организацией Японии NHK, в которой транслировалась первая встреча с гигантским кальмаром в реальном времени на пилотируемом подводном аппарате в глубоком море. у островов Огасавара.Другой был специальной выставкой под названием «Глубина», проходившей в Национальном музее природы и науки в Токио с июля по сентябрь, на которой были представлены сохранившийся образец гигантского кальмара, а также впервые снятые видеозаписи живых гигантских кальмаров. Эти два события заставили японцев, особенно молодое поколение, больше заинтересоваться гигантскими кальмарами. Такой бум внимания, направленного на гигантских кальмаров и других глубоководных существ, вероятно, увеличивает осведомленность о гигантских кальмарах среди людей в прибрежных регионах и побуждает их предупреждать средства массовой информации и местных экспертов о любых находках.
Мы также предполагаем, что Японское море может работать как большая естественная ловушка для гигантских кальмаров, мигрирующих с юга ранней весной. Их распространение широко распространяется в средние слои, которые находятся в пределах подходящей для них температурной зоны, и они продолжают расти летом и осенью. Зимой океанографические характеристики Японского моря могут вынудить гигантских кальмаров двигаться на юг и к более мелким поверхностным водам, где прибрежный водоворот Цусимского течения и сильные сезонные ветры на западе могут переносить их в прибрежные воды Японии, где они запутываются. фиксированные сети или выброшенные на берег.Для будущих исследований «гигантских кальмаров» японское побережье Японского моря в зимние месяцы будет наиболее подходящим местом для встречи с живыми гигантскими кальмарами.
Исследователи определили геном неуловимого гигантского кальмара | Генетика
Международная группа ученых секвенировала геном гигантского кальмара ( Architeuthis dux ), вида, который вдохновил поколения рассказывать сказки о легендарном Кракене. Геном Architeuthis dux позволил команде исследовать уникальные черты этого вида, включая его гигантизм и ключевую адаптацию к глубоководной среде.
Гигантский кальмар ( Architeuthis dux ), не менее 10–12 футов (3–3,7 м) в длину. Изображение предоставлено: Эди Виддер и Натан Робинсон.
Головоногие, разумная группа обитающих в океане существ, включает восьмируких осьминогов, десятируких кальмаров и каракатиц, а также наутилусов с панцирем.
Эти животные демонстрируют широкий спектр образа жизни и поведения, но, за исключением наутилусов, они характеризуются быстрым ростом и короткой продолжительностью жизни.
Они варьируются по размеру от крошечных карликовых кальмаров (средняя длина 2 см или 0,8 дюйма) до животных, которые почти на три порядка больше, таких как гигантский кальмар Architeuthis dux (средняя длина 10-12 м или 33 см). -39 футов), гигантскому кальмару Mesonychoteuthis hamiltoni (зарегистрированный вес в 500 кг делает его самым крупным из известных беспозвоночных).
Головоногие моллюски могут быстро изменять текстуру, узор, цвет и яркость своей кожи, и это позволяет создавать сложную систему связи, а также обеспечивает исключительную маскировку и мимику.Вместе они позволяют головоногим моллюскам избегать хищников и очень эффективно охотиться на добычу, что делает их одними из главных хищников в океане.
Эти замечательные приспособления головоногих моллюсков также распространяются на их геном.
«С точки зрения генов гигантский кальмар очень похож на других животных», — сказала доктор Кэролайн Альбертин, исследователь Центра регенеративной биологии и тканевой инженерии Юджина Белла Морской биологической лаборатории в Вудс-Хоул.
«Это означает, что мы можем изучать этих поистине причудливых животных, чтобы больше узнать о себе.”
Доктор Альбертин и его коллеги обнаружили, что геном Architeuthis dux имеет длину 2,7 миллиарда пар оснований ДНК — примерно 90% размера генома человека.
Они проанализировали несколько древних, хорошо известных семейств генов гигантского кальмара, проведя сравнения с четырьмя другими секвенированными видами головоногих моллюсков и геномом человека.
Они обнаружили, что важные гены развития почти у всех животных (Hox и Wnt) присутствовали в единичных копиях только в геноме Architeuthis dux .
Это означает, что это гигантское беспозвоночное существо не стало таким большим из-за дупликации всего генома — стратегии, которую эволюция потребовала давно, чтобы увеличить размер позвоночных. Итак, зная, как этот вид кальмаров стал таким гигантским, мы ждем дальнейшего изучения его генома.
Ученые также идентифицировали более 100 генов семейства протокадгеринов, которые обычно не встречаются в изобилии у беспозвоночных, в геноме Architeuthis dux .
«Считается, что протокадгерины играют важную роль в правильной работе сложного мозга», — сказал д-р.- сказал Альбертин.
«Их считали новшеством позвоночных, поэтому мы были очень удивлены, когда обнаружили более 100 из них в геноме осьминога. Это было похоже на дымящийся пистолет для того, как строить сложный мозг. И мы также обнаружили подобное расширение протокадгеринов у гигантского кальмара ».