Биоармирование это: Биоармирование лица в Минске, цены
Сеть клиник «Нефертити» в Хабаровске
Биоармирование – это коррекция возрастных изменений кожи с помощью введения биостимулирующих гелей на основе гиалуроновой кислоты. Они создают сетку из «бионитей», т.е. создают и делают более прочным внутрикожный каркас. От природы такой каркас есть у каждого из нас, но со временем волокна коллагена растягиваются, разрушаются, в результате чего появляются морщинки, теряется тургор и эластичность кожи, опущение мягких тканей лица. Биоармирование- одна из современных процедур для устранения данных проблем и профилактики дальнейших возрастных изменений кожи. Цель процедуры – лифтинг кожи и укрепление ее естественного каркаса.
История армирования берет свое начало с середины прошлого столетия, когда возникла необходимость применения методик фиксирующих кожу лица и шеи. И для этого использовались золотые (платиновые) нити. В современной косметологии чаще используют препараты на основе гиалуроновой кислоты.
Нас часто спрашивают, в чем преимущество гиалуроновой кислоты перед золотыми нитями, это:
- активная стимуляция выработки коллагена;
- высокая биологическая совместимость гиалуроновой кислоты;
- длительная стойкая увлажненность кожи;
- устранение мелких морщин , улучшение цвета, тургора кожи;
- лифтинг тканей;
- отсутствие длительной реабилитации после процедуры;
- не требуется хирургических вмешательств.

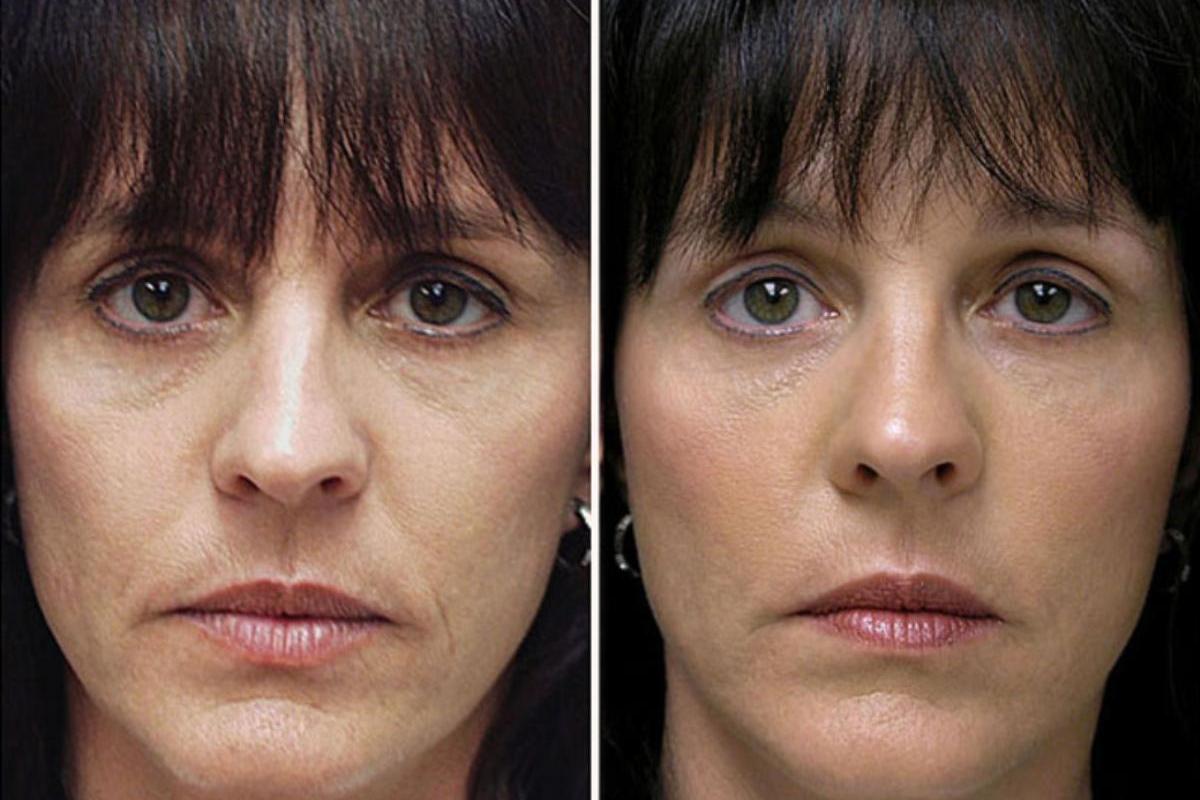
Оптимальный возраст для биоармирования 30-55 лет.
Процедура биоармирования должна проводится только квалифицированным врачом, прошедшим специальное обучение, а также в условиях процедурного кабинета с соблюдением правил асептики и антисептики.
К сожалению, очень часто женщины прибегают к услугам «кустарных» косметологов, делают процедуры на дому или в каких то других местах, где не соблюдаются санитарно-гигиенические требования, условия хранения препаратов. Да, что скрывать, иногда даже не знают , какой препарат им вводили. Тем самым подвергают опасности свою внешность, многократно увеличивая риск инфицирования кожи и других более тяжелых последствий.
В нашей клинике обязательно заполняется медицинская карта пациента, собирается подробная информация о состоянии здоровья, наличии заболеваний, аллергических реакциях, о применяемых лекарственных средствах и ранее проводимых процедурах. Затем мы подробно обсуждаем планируемую манипуляцию, выясняем , какие результаты ожидает клиент, и, конечно же, предупреждаем о всех возможных побочных эффектах.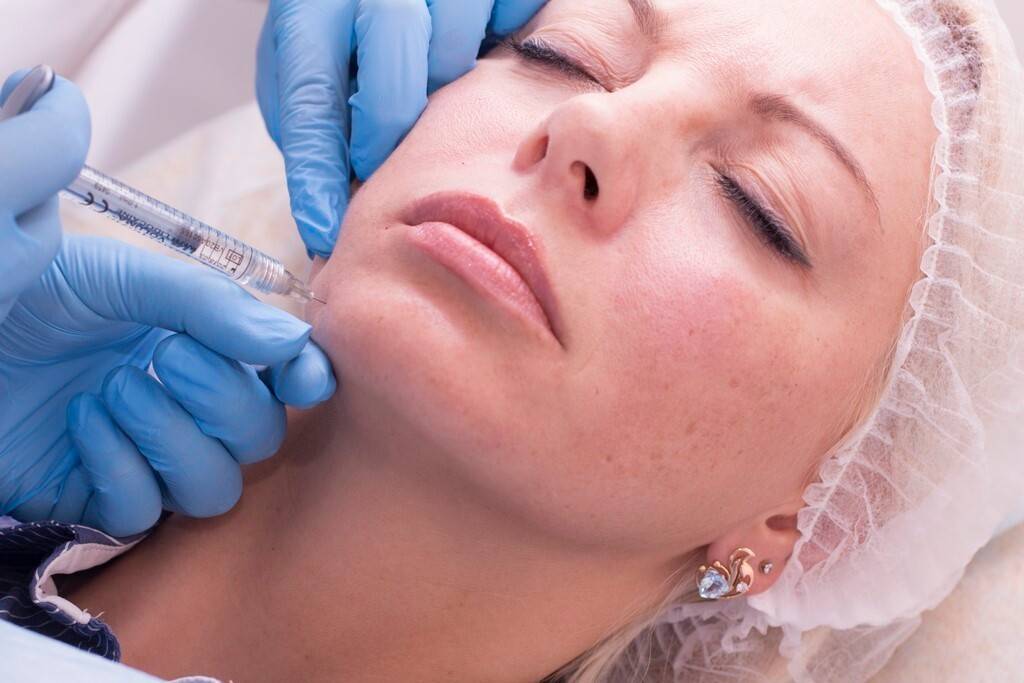 И когда решение принято, мы обязательно заполняем специальное информационное согласие на процедуру биоармирования.
И когда решение принято, мы обязательно заполняем специальное информационное согласие на процедуру биоармирования.
В клинике «Нефертити» мы используем только лучшие препараты из линеек Глайтон, Ювидерм, Белотеро. Это мировые лидеры в производстве филлеров. Они изготавливаются на крупных фармацевтических заводах, проходят тщательный контроль производства и качества, проводят крупные научные исследования эффективности и безопасности препаратов. Филлеры Глайтон, Ювидерм, Белотеро давно вошли и прочно закрепились в косметологической практике. Наши клиенты часто спрашивают, почему мы используем именно эти препараты и как мы отобрали их из представленного многообразия? На это мы с уверенностью отвечаем, что Глайтон, Ювидерм, Белотеро являются современными, высокотехнологичными филлерами, выраженный эффект которых, подтвержден данными биометрических исследований. И одно из основных условий – это высокая степень удовлетворенности наших клиентов проводимой коррекцией.
Противопоказания к процедуре биоармирования:
- нарушения свертываемости крови;
- острые заболевания, обострения хронических заболеваний, аутоиммунные заболевания;
- воспалительные процессы в местах предполагаемых инъекций;
- склонность к келоидным рубцам;
- онкологические заболевания;
- беременность и период лактации.

Перед введением кожа тщательно очищается и обрабатывается антисептическим раствором. Затем наносится анестезирующее средство на 20-30 минут. Препарат вводится по определенной схеме, небольшими порциями. После процедуры отмечается небольшой отек и покраснение кожи, которые чаще всего проходят в течение суток. В зависимости от состояния сосудов кожи могут быть небольшие синячки – в таком случае мы назначаем на дом заживляющие, рассасывающие средства для местного использования.
Видимый, стойкий эффект наступает через 10-14 дней и зависит от начального состояния кожи и количества процедур.
После процедуры существуют некоторые ограничения, которых необходимо придерживаться, дабы избежать побочных эффектов:
- воздержаться от посещения бань, саун, соляриев в течение двух недель;
- не использовать никакие косметические средства в местах инъекций в течение 12 часов;
- воздержаться от массажей лица около 2х месяцев.
Таким образом, процедура биоармирование заняла достойное место в anti-age терапии.
Биоармирование гелями в Санкт-Петербурге – цены на процедуру, отзывы
Биоармирование — это омоложение кожи лица путем введения специальных гелей (Ювидерм, Радиес и др.) на основе гиалуроновой кислоты, который образует «паутинку» из бионитей, т.е. создает подкожный «каркас». Такой каркас существует от природы у каждого человека, но со временем волокна коллагена растягиваются, в результате лицо начинает терять контур. Биоармирование — это один из современных способов борьбы со старением кожи лица и шеи
Кому показано биоармирование:
Мы советуем всем пациентам. Особенно эффективно сочетание мезотерапии гиалуроновой кислотой и биоармирования. Для проведения процедуры биоармирования самым оптимальным возрастом считается 35-50 лет.
Что ожидать от процедуры биоармирования:
Биоармирование не только восполнит недостаток гиалуроновой кислоты в коже, но и позволит коже обрести собственный эластичный «каркас», который избавит от существующих морщин и будет надежно предохранять от возникновения новых. Кроме того, кожа становится необычайно эластичной и упругой, улучшается ее цвет, контур лица становится более четким. Биопрепарат имеет малый вес, поэтому не отяжеляет кожу. Это очень важно, ведь именно гравитационный птоз является одним из самых важных факторов, приводящих к старению мягких тканей. Таким образом, биоармирование стоит между косметологическими средствами и оперативным вмешательством, взяв все лучшее — безопасность и биологичность препаратов и сильный эффект, аналогичный результатам пластической хирургии. Омоложение лица в данном случае происходит максимально физиологично и надежно.
Кроме того, кожа становится необычайно эластичной и упругой, улучшается ее цвет, контур лица становится более четким. Биопрепарат имеет малый вес, поэтому не отяжеляет кожу. Это очень важно, ведь именно гравитационный птоз является одним из самых важных факторов, приводящих к старению мягких тканей. Таким образом, биоармирование стоит между косметологическими средствами и оперативным вмешательством, взяв все лучшее — безопасность и биологичность препаратов и сильный эффект, аналогичный результатам пластической хирургии. Омоложение лица в данном случае происходит максимально физиологично и надежно.
Как проходит процедура:
При биоармировании с помощью инъекций кожа не травмируется, и не требуется последующего реабилитационного периода. После процедуры биоармирования на коже остаются незначительные следы от уколов, которые быстро исчезают. При применении врачом обезболивающего крема инъекции не доставляют болезненных ощущений.
Что это такое биоармирование и кому оно назначается?
Биоармирование – это восполнение общего контура лица.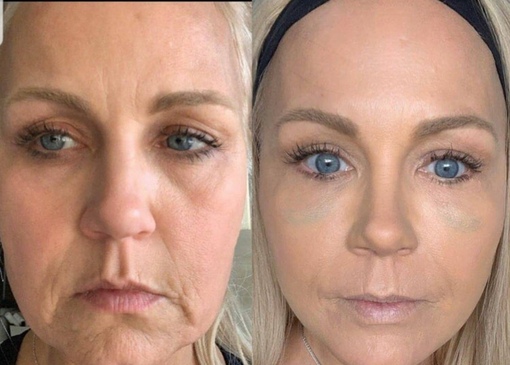 Было доказано учеными, что, к сожалению, при достижении определенного возраста земное притяжение, гравитация отрицательно влияет на перемещение объема жировых пакетов, которые у нас есть в области щек, очень отрицательно на них сказывается. Биоармирование восполняет данный контур, придаёт чёткость контуру лица, в косметологии даже есть термин – лицо в виде сердечка. Им всегда отличалась актриса Анжелина Джоли, которая является эталоном на протяжении уже десятилетия в своей красоте. Что такое сердечко? Это более чётко выраженные объемы в верхней трети лица и меньшее содержание объемов в нижней трети лица. Именно в биоармировании идет применение тех же препаратов контурной пластики – высокомолекулярной гиалуроновой кислоты, которая восполняет структуру нашего лица.
Было доказано учеными, что, к сожалению, при достижении определенного возраста земное притяжение, гравитация отрицательно влияет на перемещение объема жировых пакетов, которые у нас есть в области щек, очень отрицательно на них сказывается. Биоармирование восполняет данный контур, придаёт чёткость контуру лица, в косметологии даже есть термин – лицо в виде сердечка. Им всегда отличалась актриса Анжелина Джоли, которая является эталоном на протяжении уже десятилетия в своей красоте. Что такое сердечко? Это более чётко выраженные объемы в верхней трети лица и меньшее содержание объемов в нижней трети лица. Именно в биоармировании идет применение тех же препаратов контурной пластики – высокомолекулярной гиалуроновой кислоты, которая восполняет структуру нашего лица.
Можно ли убрать объём? Есть обратная история?
– Это касается вопроса об увеличении объёма губ. Говорят про женщину: «Закачала себе и ходит со своими губищами». Женщину убедили, что она действительно переборщила. Можно ли убрать лишнее?
Можно ли убрать лишнее?
На сегодняшний день тот перебор в размерах губ, который был моден еще 4-5 лет назад, неактуален. Действительно, было поветрие, что у всех должны быть крупные губы. У любой женщины губы должны быть чувственными, выраженными. Сейчас тенденции совершенно другие, приветствуются губы немного припухлые, чтобы никто даже не подумал, что вы сделали процедуру у врача-косметолога.
Вывести можно, я сталкивалась с такой проблематикой, когда приходили пациентки с очень большими губами. Первым делом я спрашивала, что было введено. Раньше губы могли увеличить биополимером. К счастью, этот материал запрещен к применению законодательством Российской Федерации, но до сих пор есть умельцы, которые подпольно могут вводить эти препараты. Данный препарат, кроме как хирургическим путем, никак не выводится из организма. Поэтому данную пациентку можно послать только к хирургу.
Почему пациентки обращаются к биополимерам, если препарат запрещен? Какие негативные последствия от него?
Биополимер может мигрировать по всему телу. Данный полимер, введенный в область губ, могли находить в области шеи, плеча, даже в области дельтовидной мышцы, то есть, могли быть совершенно разные места. Материал полностью чужеродный для организма, полная синтетика. Естественно, организм начинает его отторгать. Я могу объяснить, из-за чего обращаются. Иногда чисто из-за лени. Препарат с гиалуроновой кислотой, который мы вводим в губы, исчезает, деградируется из организма через какое-то время, а биополимер можно один раз ввести, и больше не обращаться к врачу-дерматокосметологу для корректировки и восполнения потерянного объема.
Данный полимер, введенный в область губ, могли находить в области шеи, плеча, даже в области дельтовидной мышцы, то есть, могли быть совершенно разные места. Материал полностью чужеродный для организма, полная синтетика. Естественно, организм начинает его отторгать. Я могу объяснить, из-за чего обращаются. Иногда чисто из-за лени. Препарат с гиалуроновой кислотой, который мы вводим в губы, исчезает, деградируется из организма через какое-то время, а биополимер можно один раз ввести, и больше не обращаться к врачу-дерматокосметологу для корректировки и восполнения потерянного объема.
Биоармирование и биорепарация (FACE-лифтинг) — Клиническая больница «РЖД-Медицина» города Нижний Новгород
| КОСМЕТОЛОГИЯ | 8 986 760 16 68 |
Биоармирование – это активный синтез коллагеновых и эластиновых волокон с помощью нано технологий определяющий биологический face-лифтинг. Это процедура введения в кожу биостимулятора, который глубоко в дерме образует паутинку из бионитей. Безопасно, гипоалергенно.
Это процедура введения в кожу биостимулятора, который глубоко в дерме образует паутинку из бионитей. Безопасно, гипоалергенно.
Биорепарация – новая концепция в эстетической медицине, направленная на предотвращение старения и омоложение кожи лица. Омолаживающее действие заключается именно в физиологическом восстановлении объемов с совместным и одновременным ревитализирующим действием, которое заключается в стимуляции к обновлению и реструктуризации тканей.
Ход процедуры:
Поверхностные слои кожи прошиваются препаратом с гиалуроновой кислотой по специальной технике с целью создания каркаса и уплотнения кожи, разглаживания морщин и получения лифтинга. Лицо обрабатывается зонально. После процедуры возможны гематомы, отёчность в месте введения препарата от 2 до 5 суток. Возможна аппликационная анестезия. Лечение рубцовой ткани.
Результат: дермальный (кожный) лифтинг, уплотнение и разглаживание кожи.
| Наименование препарата | Стоимость, рубли |
| HYALREPAIR 02 1,5мл | 7 000 |
| Apriline 1,0 мл | 10 900 |
| HYALREPAIR 04 /08 1,5 мл | 7 000 |
| Jalupro 3,0 мл | 6 500 |
| Jalupro HMV | 8 500 |
| Meso-Wharton P 199 1,5 мл | 10 000 |
| Meso-Xanthin F 199 1,5 мл | 10 000 |
| Meso-Sculpt C71 1 мл | 10 000 |
| MESOEYE C 71 1 мл | 10 000 |
| Коллост-гель 7% 1.5мл | 1 шприц 10 402 |
| Коллост-гель 7% 1 мл | 1 шприц 8 600 |
| Коллост-гель 15% 1.5мл | 1 шприц 16 500 |
| Анестезия инъекционная местная | 550 |
| Анестезия безинъекционная | 668 |
| Анестезия методом нанесение геля “МИОРЕЛАКСАНТ” | 700 |
| Гель Мио релаксант ( анестезия) | 400 |
цены на биоармирование лица в клинике Оксфорд Медикал
Многие годы в борьбе с неприятными возрастными изменениями кожи самым популярным и наиболее эффективным был метод круговой подтяжки лица. Однако это довольно сложная операция с определенными рисками и долгим реабилитационным периодом. На смену данной методике какое-то время назад пришла прошивка золотыми нитями. Данная процедура также имеет недостатки: нити из чужеродного для организма материала остаются в коже пациента навсегда, что в будущем ограничивает либо полностью исключает возможность применения других методик омоложения.
Однако это довольно сложная операция с определенными рисками и долгим реабилитационным периодом. На смену данной методике какое-то время назад пришла прошивка золотыми нитями. Данная процедура также имеет недостатки: нити из чужеродного для организма материала остаются в коже пациента навсегда, что в будущем ограничивает либо полностью исключает возможность применения других методик омоложения.
Процедура биоармирования мезонитями, применяемая в клинике Оксфорд Медикал, появилась сравнительно недавно. На сегодняшний момент это настоящая альтернатива хирургическим пластическим операциям. Биоармирование мезонитями всего за одну процедуру безопасно устраняет морщины и отлично омолаживает лицо на несколько лет.
ДЛЯ ЧЕГО ПРИМЕНЯЕТСЯ БИОАРМИРОВАНИЕ МЕЗОНИТЯМИ
Процедура успешно борется с:
- дряблой и обвисшей кожей на лице;
- глубокими морщинами на лбу;
- кисетными морщинами;
- носогубными складками;
- опущением глаз, бровей и уголков губ.
Биоармирование — очень эффективный современный способ возвратить лицу его ровные и красивые очертания.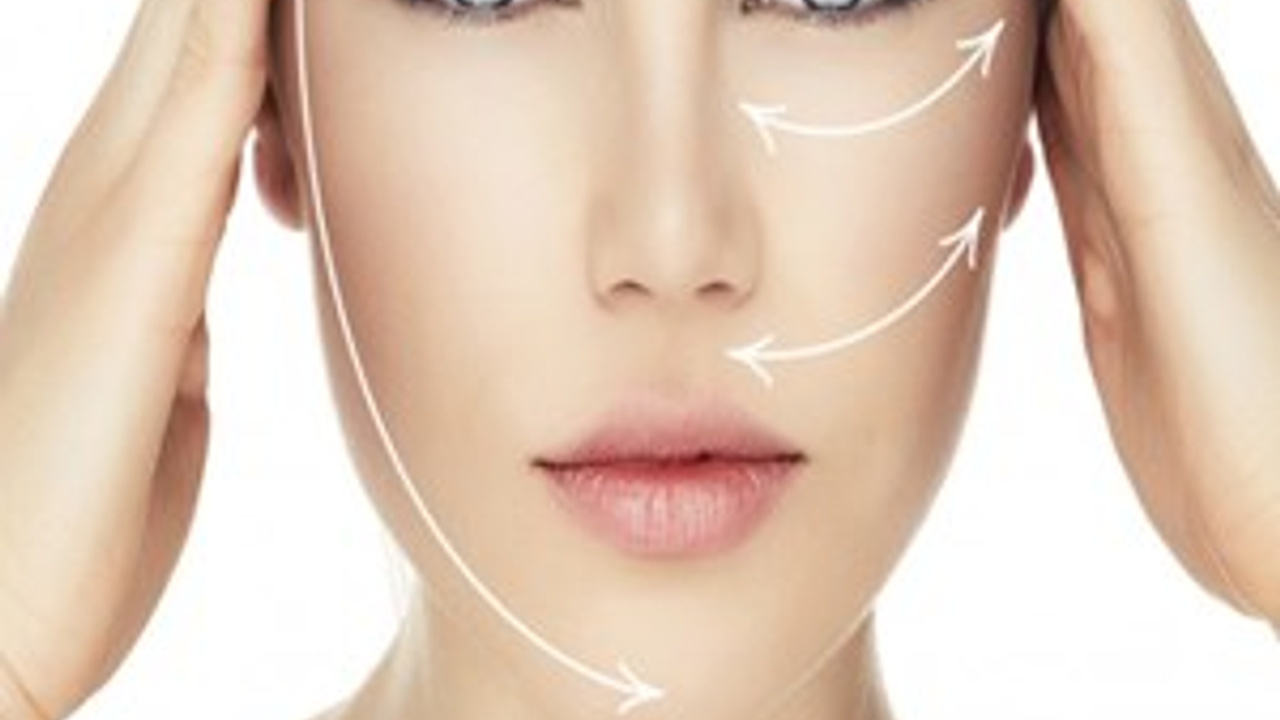 Во время процедуры в области лица и шеи под кожу вводится специальный биостимулятор, который интенсивно воздействует на клетки, вырабатывающие в организме белки, эластин и коллаген. Клетки в процессе армирования лица активно обновляются и растут. Результатом процедуры является упругая кожа, разглаженные морщины и свежий цвет лица.
Во время процедуры в области лица и шеи под кожу вводится специальный биостимулятор, который интенсивно воздействует на клетки, вырабатывающие в организме белки, эластин и коллаген. Клетки в процессе армирования лица активно обновляются и растут. Результатом процедуры является упругая кожа, разглаженные морщины и свежий цвет лица.
Биоармирование — это золотая середина между традиционными косметологическими средствами и пластической хирургией. Данная процедура взяла от этих средств все наилучшее — натуральность и безопасность консервативных методов и высокую результативность оперативного вмешательства.
ПРЕИМУЩЕСТВА МЕТОДИКИ
- максимально физиологичная, безопасная и надежная коррекция морщин;
- длительность процедуры — до 40 минут;
- биоармирование проводится без анестезии или с минимальным обезболиванием;
- не требуется время для реабилитации;
- техника биоармирования хорошо сочетается с методом контурной пластики.
Биоармирование мезонитями имеет некоторые противопоказания:
- онкологические заболевания;
- расстройства нервной системы;
- период беременности и кормления грудью;
- воспалительные заболевания кожи;
- наличие инъекционных имплантатов.
ПОЧЕМУ КЛИНИКА «ОКСФОРД МЕДИКАЛ»
- Процедуру биоармирования мезонитями рекомендуется проводить в крупном медицинском центре, каковым и является клиника «Оксфорд Медикал».
- Для получения максимального лифтинг-эффекта, специалист, проводящий процедуру, обязан в идеале знать анатомию и в совершенстве владеть техникой введения мезонитей.
- Одно-единственное неточное движение может спровоцировать появление новых складок, исправить которые потом довольно сложно.
- Колоссальный опыт и абсолютный профессионализм — основной капитал специалистов «Оксфорд Медикал».
Биоармирование лица (кожи) в Казани – «Эстетик Сити»
Биоармирование — это физиологичный и надёжный способ придать лицу первоначальные контуры. В области лица и шеи под кожу вводится сильный биостимулятор — это специальное, гелеобразное вещество. Оно чрезвычайно интенсивно воздействует на все клетки, которые вырабатывают белки, эластин и коллаген, совершая титаническую работу, название которой – коррекция морщин. Клетки во время армирования лица активно обновляются и растут. В результате процедуры кожа становится очень упругой, эффективно разглаживаются морщины и значительно улучшается цвет.
Клетки во время армирования лица активно обновляются и растут. В результате процедуры кожа становится очень упругой, эффективно разглаживаются морщины и значительно улучшается цвет.
Секрет такого эффекта – в использовании для армирования тела натуральной или синтезированной гиалуроновой кислоты, которая имеет первостепенное значение в сохранении молодости и красоты. Биоармирование занимает выгодную позицию между консервативными косметологическими средствами и оперативным вмешательством, взяв от этих средств все лучшее – натуральность и безопасность традиционных препаратов и сильный эффект, аналогичный результатам пластической хирургии. Коррекция морщин в данном случае происходит максимально физиологично и надежно.
Показания к процедуре:
Био армирование тела активно помогает:
- Произвести коррекцию морщин;
- Укрепить тургор кожи;
- Скорректировать овал лица.
Для проведения процедуры армирования тела и лица самым оптимальным возрастом считается 35-50лет. Гораздо сложнее возобновить процесс восстановления кожного покрова после 50 лет, поэтому эффективность, которую дает биоармирование, резко падает. Обычно биоармирование востребовано, если снижена упругость кожи, что приводит к обвисанию лица и оплыванию его контуров. В таком случае коррекция морщин, проведенная с помощью биоармирования, даст превосходные результаты.
Гораздо сложнее возобновить процесс восстановления кожного покрова после 50 лет, поэтому эффективность, которую дает биоармирование, резко падает. Обычно биоармирование востребовано, если снижена упругость кожи, что приводит к обвисанию лица и оплыванию его контуров. В таком случае коррекция морщин, проведенная с помощью биоармирования, даст превосходные результаты.
Пройдите простой тест и узнайте, показана ли Вам процедура армирования:
Подойдите к зеркалу и наклоните голову. Если во время наклона головы сместился овал лица, появились складки, то Вам стоит пройти процедуру армирования биогелем. Биоармирование не только поможет «вернуть на место» лицо, но также заметно омолодит кожу, сделав ее более свежей и упругой. Кроме того, проведя биоармирование, Вы надолго забудете о проблеме старения: введенный биогель будет постоянно помогать коже обновляться и омолаживаться – коррекция морщин будет действовать довольно долго.
ПРОТИВОПОКАЗАНИЯ
- Приём антикоагулянтов;
- Нарушения свёртываемости крови;
- Любые заболевания, которые находятся в стадии обострения;
- Инфекционный и/или воспалительный процесс в тех местах, где будет необходимо производить инъекции;
- Повышенная склонность к образованию келоидных рубцов на коже;
- Беременность, а так же период грудного вскармливания.
Еще до проведения коррекции морщин стоит обговорить все эти моменты с косметологом.
Как проходит процедура:
Перед тем, как провести сеанс биоармирования, врач-дерматолог собирает всю необходимую информацию о состоянии здоровья пациента и о его типе кожи. Далее провидится сама процедура биоармирования. По окончании всех манипуляций на коже остаются едва заметные следы от уколов, которые бесследно исчезнут через день. Во время процедуры применяется специальный обезболивающий крем, поэтому инъекции безболезненны. Биоармирование проводится в амбулаторных условиях и не требует реабилитационного периода. В отличие от оперативного вмешательства, биоармирование – относительно простая процедура, гарантирующая великолепный эффект при минимальных возможных последствиях.
Услуга оказывается в следующих Центрах красоты и здоровья:Записатьcя
Биоармирование лица в Спб — цены и отзывы на Биоармирование лица Санкт-Петербург
Биоармирование это процедура, основанная на технике армирования золотыми нитями, только вместо нитей под кожу вводится специальный гель, который восполняет недостаток гиалуроновой кислоты и стимулирует выработку коллагена. Создаваемый таким образом каркас из новой соединительной ткани восстанавливает контуры лица и омолаживает кожу, возвращая ей эластичность.
По истечении времени введенный гель рассасывается, но эффект сохраняется на длительный срок. Биоармирование хорошо сочетается с мезотерапией и биоревитализацией, гарантируя впечатляющие результаты, которые раньше могла обеспечить только пластическая хирургия.
Кому подойдет биоармирование лица?
Данная процедура подходит для людей от 35 лет, когда начинают появляться первые признаки старения: опущение уголков рта, нарушение овала лица и углубление мимических морщин.
Однако, при более выраженных изменениях, которые наступают после 50 лет, биоармирование овала лица не поможет, но придаст коже упругости и сияния.
Преимущества процедуры
Биоармирование имеет значительные преимущества по сравнению с другими методами омоложения.
Отсутствие хирургического вмешательства. Биогель вводится под кожу при помощи тончайших игл, в отличие от армирования золотыми нитями, введение которых производится хирургическим путем.
Безопасность. Гиалуроновая и полимолочная кислота отлично усваиваются организмом, в отличие от нитей и препаратов с химическим составом.
Реабилитация. Период реабилитации проходит быстро и не требует нахождения в стационаре.
Быстрый результат. Результат процедуры становится заметен через 1-2 недели.
Противопоказания
Обратите внимание, биоармирование лица имеет несколько противопоказаний, с которыми стоит ознакомиться перед процедурой:
- Наличие заболеваний иммунной системы;
- Нарушение свертываемости крови, или прием препаратов, которые влияют на свертываемость крови;
- Склонность организма к появлению рубцов. Так как препарат вводится под кожу с помощью игл, то в местах введения могут остаться рубцы;
- Наличие избытка кожи. Если у вас много лишней кожи на лице, то данная процедура не сможет решить эту проблему.
Техника проведения процедуры
Биоармирование лица проводится курсами по 3-4 процедуры с интервалом 1-1.5 месяца. Один сеанс длится около часа.
Во время сеанса гель вводится в кожу параллельно ее поверхности серией инъекций с помощью тончайших игл (векторный лифтинг) или гибких канюль (софт-лифтинг).
Особенность софт-лифтинга состоит в том, что канюли имеют закругленное окончание, которое раздвигает ткани, не травмируя их. Это позволяет исключить вероятность возникновения синяков и кровоподтеков.
Перед проведением процедуры определяется индивидуальный болевой порог и наносится анестезирующий крем. После того, как крем подействовал, можно приступать к процедуре. Препарат вводится под кожу по специальной схеме.
Введение геля производится по специальной технике: гель не вкачивается, а остается в коже по мере медленного извлечения иглы или канюли. В результате область коррекции «прошивается» тонкими линиями препарата. Направление этих линий зависит от тех задач, которые решает косметолог для каждого конкретного пациента.
Созданная в результате инъекций «прошивка» является основой для будущего коллагенового каркаса, который будет развиваться под действием активного компонента.
Имеет значение, какое вещество является основой вводимого геля:
- Гиалуроновая кислота. Применяется для увлажнения кожи. Препарат действует на верхние слои кожи, восполняя недостаток кислоты в клетках, увлажняя кожу и восстанавливая внешний вид и здоровый цвет. Эффект от процедуры длится до 6 месяцев.
- Полимолочная кислота. Данный препарат применяется для более глубокого восстановления при ярко-выраженных морщинах и опущении овала лица. После введения препарата, он стимулирует выработку коллагена в тканях и питает клетки, восстанавливая каркас лица. Кожа приобретает прежнюю эластичность и подтягивается, разглаживая морщины. Улучшается ее общее состояние. Полимолочная кислота дает лучший результат, а эффект сохраняется до 3 лет.
Выбор препарата определяется в ходе консультаций с врачом-косметологом на основании пожеланий пациента и индивидуальных особенностей его организма.
18.3A: Гибридные зоны — Биология LibreTexts
Со временем два вида могут еще больше расходиться или воссоединяться, в зависимости от силы приспособленности и репродуктивных барьеров гибридов.
Повторное подключение после спецификации
Видообразование происходит в течение эволюционного периода. Когда возникает новый вид, наступает переходный период, в течение которого близкородственные виды продолжают взаимодействовать.
После видообразования два вида могут рекомбинировать или даже продолжать взаимодействовать бесконечно.Отдельные организмы будут спариваться с любыми находящимися поблизости особями, с которыми они способны к размножению. Область, где два близкородственных вида продолжают взаимодействовать и размножаться, образуя гибриды, называется гибридной зоной. Со временем гибридная зона может измениться в зависимости от силы приспособленности и репродуктивных барьеров гибридов.
Рисунок \ (\ PageIndex {1} \): Видообразование и гибридная зона : После того, как произошло видообразование, два отдельных, но тесно связанных вида могут продолжать производить потомство в области, называемой гибридной зоной.В зависимости от репродуктивных барьеров и относительной приспособленности гибридов может произойти усиление, слияние или стабильность.
Гибриды могут иметь меньшую приспособленность, большую приспособленность или примерно такой же уровень приспособленности, как и чистокровные родители. Обычно гибриды менее приспособлены; следовательно, воспроизводство для получения гибридов со временем будет уменьшаться, что подталкивает два вида к дальнейшему расхождению в процессе, называемом подкреплением. Этот термин используется, потому что низкий успех гибридов усиливает первоначальное видообразование.Если гибриды менее приспособлены, чем родители, происходит усиление видообразования, и виды будут продолжать расходиться до тех пор, пока они не перестанут спариваться и производить жизнеспособное потомство.
Если гибриды подходят или лучше, чем родители, или репродуктивные барьеры ослабевают, два вида могут снова слиться в один вид (воссоединение). Чтобы гибридная форма сохранялась, она, как правило, должна уметь использовать имеющиеся ресурсы лучше, чем любой родительский вид, с которыми в большинстве случаев ей придется конкурировать.
Со временем, посредством процесса, называемого гибридным видообразованием, сами гибриды могут стать отдельным видом. Когда-то считалось, что добиться репродуктивной изоляции между гибридами и их родителями особенно сложно; таким образом, гибридные виды считались чрезвычайно редкими. Поскольку в 1990-х годах анализ ДНК стал более доступным, гибридное видообразование оказалось довольно распространенным явлением, особенно у растений.
Ученые также заметили, что иногда два вида остаются отдельными, но продолжают взаимодействовать с образованием нескольких гибридных особей; это классифицируется как стабильность, поскольку реальных чистых изменений не происходит.Чтобы гибридная зона была стабильной, потомство, полученное от гибридов, должно быть менее приспособленным, чем представители родительского вида.
Выборочный обзор эволюции со времен Дарвина — первые 150 лет. Под редакцией М. А. Белла, Д. Дж. Футуямы, В. Ф. Эанеса и Дж. С. Левинтона
J Exp Anal Behav. 2012 Март; 97 (2): 249–260.
Массачусетский университет в Амхерсте
Для переписки можно обращаться к Джону Донахью, почетному профессору факультета психологии, Программа поведенческой нейробиологии, Массачусетский университет, Амхерст, Массачусетс 01003 (электронная почта: ude.ssamu.hcysp@eohanodj)
Поступила 21 октября 2011 г .; Принято 16 декабря 2011 г.
Copyright Society for the Experimental Analysis of Behavior, Inc.
Abstract
Этот обзор фокусируется на параллелях между селекционистскими науками эволюционной биологии и анализом поведения. В селекционизме сложные явления интерпретируются как совокупные продукты относительно простых процессов, действующих во времени — естественного отбора в эволюционной биологии и подкрепления в анализе поведения. Поскольку эволюционная биология является более зрелой наукой, изучение факторов, приведших к победе естественного отбора, дает ключи к разгадке, посредством которых подкрепление может постигнуть аналогичную судьбу в науке о поведении.
Ключевые слова: селекционизм, эволюция, естественный отбор, подкрепление, суеверие, молярно-молекулярная проблема, генетика
Эволюция через естественный отбор, возможно, является главным научным достижением 19 века. Но почему при анализе поведения следует уделять особое внимание изменениям в эволюционной биологии за 150 лет после публикации книги Дарвина «О моделях O видов »? Ответ заключается в том, что анализ поведения — уникальное явление для наук о поведении — также придерживается селекционного подхода к сложности (Donahoe, 1983; 2003; Palmer & Donahoe, 1992).В селекционистском подходе сложные явления рассматриваются как совокупный продукт относительно простых процессов, действующих во времени. Для Дарвина относительно простым процессом был естественный отбор, при котором дифференцированный репродуктивный успех позволял накапливать наследственные различия. Накопление этих различий позволило им стать непропорционально представленными в последующих поколениях группы межпородных организмов. Для анализа поведения относительно простым процессом является подкрепление, которое по-разному способствует тому поведению, за которым следуют определенные события, называемые подкреплением.В результате продукты отбора путем подкрепления все чаще представлены в поведенческом репертуаре отдельного организма. В настоящем обзоре исследуется тема, согласно которой история более зрелой селекционистской науки эволюционной биологии дает ключи к анализу поведения в его попытках понять сложное поведение.
После краткого описания общих характеристик процессов отбора этот обзор состоит из двух основных разделов. В первом разделе исследуются параллели между естественным отбором и отбором по подкреплению.Второй раздел идентифицирует несколько неожиданный аспект Evolution Since Darwin ( ESD ) и рассматривает его значение для будущего анализа поведения.
Из-за целенаправленности настоящего обзора ряду важных тем в этой монографии из 39 авторов, состоящей из 22 глав, не уделяется должного внимания. Я упоминаю здесь только два. Первый посвящен истории эволюционной биологии (см. Kokkop & Jennions, стр. 293). (Условие, используемое здесь для ссылки на работу в ESD , состоит в том, чтобы указать имя автора с последующим соответствующим номером страницы.Глава Боулера размышляет о том, каким было бы нынешнее состояние эволюционной биологии, если бы генетические механизмы наследственности были бы открыты с до , принцип естественного отбора. (Книга Боулера Затмение Дарвина , 1983, пересчитывает годы между публикацией Происхождение видов и принятием естественного отбора в биологии, и известна читателям JEAB ; Катания, 1987). Боулер провокационно заключает, что «… мир без Дарвина… это что-то, что было бы удивительно похоже на крайность (т.е., неселективистская) версия современной эволюционной биологии развития »(с. 43). То есть, вместо функционального отношения к сложности, поощряемого естественным отбором, эволюционная биология приобрела бы структуралистский оттенок, сосредотачивающийся в первую очередь на механистических деталях развития и морфологии. Вторая тема, которая не включена в настоящий обзор, — это эволюция человека. В главе Уайта дается обзор палеонтологических исследований происхождения человека, который опровергает некоторые утверждения о том, что летопись окаменелостей эволюции человека нехарактерно скудна.Фактически, изучение происхождения человека является чрезвычайно активным и продуктивным направлением текущей работы как на палеонтологическом уровне (например, Carlson et al., 2011), так и на молекулярном уровне (например, Edinborval et al., 2011; Noonan et al. ., 2006; Йотова и др., 2011). Например, много было сделано из открытия, что люди и шимпанзе имеют примерно 98% общих структурных генов (то есть генов, кодирующих белок). Однако регуляторные гены, которые управляют действием структурных генов, сильно отличаются от шимпанзе и гораздо менее изменчивы у людей, чем у других видов (King & Wilson, 1975; Kirkpatrick, p.181ff). Кроме того, само количество генов не связано со сложностью: геном человека содержит около 20 000 генов, тогда как геном риса содержит более 40 000 генов. Наш вид, по-видимому, является продуктом естественного отбора очень ограниченной комбинации относительно небольшого числа генов (Wray, стр. 234). Хотя генетический состав людей и других видов становится все более известным, повторяющаяся тема в ESD заключается в том, что связь между геномом и фенотипом остается нерешенной загадкой.Как выразились некоторые участники, «… молекулярно-генетическая основа сложности организма остается в значительной степени необъясненной» (Чжан, стр. 97), «определение фенотипических эффектов на основе нуклеотидных изменений остается сложной задачей» (Шулер, стр. 283) и «по сравнению с нашими знаниями. геномов наши знания о фенотипах остаются поверхностными »(Hoekstra, p. 642). Наконец, я хотел бы отметить, что ESD всегда хорошо написаны и тщательно отредактированы.
ПРИРОДА ПРОЦЕССОВ ОТБОРА
Дарвинизм, или селекционизм , как более широко известен подход, дает представление о происхождении сложности, которое не влечет за собой принципы, которые непосредственно навязывают сложность (Richerson & R.Бойд, стр. 361). Напротив, сложность является возможным — хотя и не неизбежным — результатом повторяющегося действия процессов низшего порядка. Как общий подход, потенциальный диапазон селекционизма простирается далеко за пределы эволюционной биологии. Любые ограничения селекционизма проистекают в первую очередь из дотеоретических обязательств — особенно из вида имплицитного рационализма, который пронизывает многие описания человеческого поведения, как, например, в лингвистике и когнитивной психологии (Donahoe, 1983). Селекционизм был продуктивно воплощен в таких разнообразных областях, как проектирование электронных схем (Голландия, 1975; 1992), экономика (Харфорд, 2011) и космология (Смолин, 1997).
Процесс отбора состоит из трех взаимозависимых фаз — вариации, отбора и удержания. Вариант обеспечивает исходный материал, на котором действует отбор. Это источник любой новизны, возникающей в результате повторяющихся циклов процесса отбора. Отбор действует только на уже существующие варианты. Это четко признается в эволюционной биологии, где Уэйкли в ESD комментирует, что «естественный отбор может происходить только в том случае, если особи вида различаются» (стр.119). Используя термин Дональда Кэмпбелла, вариация является «ненаправленной» (Campbell, 1974). То есть изменение происходит независимо от фактора выбора. Второй этап процесса отбора — это сам отбор. Отбор потенциально приводит к сложности, когда какое-то событие, действующее на популяцию вариантов, отдает предпочтение (или отрицает) один вариант по сравнению с другим. Отбор придает процессу иллюзию цели, когда случайные обстоятельства выбора остаются постоянными или медленно меняются с течением времени. Процессы отбора не имеют телеологической направленности.Будущее не тянет настоящее к себе; вместо этого прошлое толкает настоящее в будущее. Траектория отбора полностью зависит от относительного постоянства случайностей выбора и совокупности вариантов. Будущие непредвиденные обстоятельства «ожидаются» только постольку, поскольку они встречались в прошлом. Удержание — это третий этап процесса отбора. Сохранение позволяет выбранным вариантам сохраняться достаточно долго, чтобы они внесли свой вклад в изменение, на которое будут действовать будущие выборы.Без сохранения выборки не могут накапливаться, и даже возможность сложности исключается. (Обсуждение последствий селекционизма для поведения см. В Catania, 1995; Donahoe & Wessells, 1980; Donahoe & Palmer, 1994; Palmer & Donahoe, 1992; Skinner, 1953; 1966; и Staddon & Ettinger, 1989. философские подходы, см. Dennett, 1995; Gayon, 1992; Hull, 1973; Mayr, 1988; и Sober, 1984). естественный отбор.Например: «Этот поток генетических данных будет мало полезен для биологов-эволюционистов, если экологический контекст места организма в окружающей среде не будет хорошо изучен (МакПик, стр. 345). Или, опять же, «изменение морфологии животных часто предшествует изменению в поведении» и «связи между генами, нейронными цепями и эволюция сложного и адаптивного поведения остаются главным рубежом в биологии» (Hoekstra, pp. 645–645). 646). Сам Дарвин одобрительно цитируется с аналогичным эффектом: «Переход в новую нишу или адаптивную зону, почти без исключения, инициируется изменением поведения» (цит. По: Mayr, 1963, p.604). Однако, несмотря на эти признания, в ESD практически ничего не говорится о научном изучении поведения. Ватсон и бихевиоризм кратко упоминаются как обеспечивающие, что «поведенческие исследования становились все более количественными и менее антропоморфными», и как обеспечивающие «первые инструменты для выявления и количественной оценки поведенческих черт» (Kokko & Jennions, p. 293). Но ни Уотсон, ни бихевиоризм не фигурируют в обширном индексе, а Скиннер и другие бихевиористы полностью игнорируются, хотя существует значительная литература о параллелях между отбором путем подкрепления и естественным отбором.Фактически, некоторые участники ESD ошибочно — если это понятно — повторяют широко распространенную, но ошибочную точку зрения (например, Simon, 1980) об «ограничениях [поведенческого подхода] при работе с видами, для которых теория разума кажется необходимым »(Kokko & Jennions, стр. 293). Читая этот комментарий, студенты Скиннера могут утешиться следующим утверждением о взглядах Дарвина: «Последующие исследования эволюции человека часто приписывают Дарвину идеи, которые на самом деле не могут быть найдены в его трудах, а только во второстепенных, третичных или даже более производных. и менее точные источники »(Уайт, с.522).
Вариант
Прежде чем исследовать параллели между тремя фазами процесса отбора при естественном отборе и отборе путем подкрепления, должно быть очевидно, что параллели являются функциональными, а не структурными. То есть физические события и механизмы, реализующие три фазы, различаются, но эффекты фаз концептуально связаны. В случае естественного отбора вариация относится к различиям между характеристиками (фенотипом) различных организмов в племенной популяции.Популяция вариантов выбора поведения принципиально различна. Здесь вариация относится к различиям внутри популяции в поведении (поведенческом репертуаре) одного организма . Различие между популяциями, над которыми действует отбор, — вот что мотивирует методологическое расхождение между изучением поведения отдельных организмов в анализе поведения и групп организмов в нормативной психологии. Нормативная психология использует групповые методологии, которые были разработаны для изучения естественного отбора, а не индивидуальные методологии, подходящие для отбора путем подкрепления (см.Сидман, 1960). В тех случаях, когда поведенческие аналитики используют групповые методологии, обычно они сообщают о своих выводах таким образом, чтобы более эффективно взаимодействовать с другими поведенческими дисциплинами.
Вариации в эволюционной биологии
Открытие генетической основы наследования более чем через 40 лет после О происхождении видов привело к способности идентифицировать различия между людьми на уровне наблюдения ниже фенотипа, а именно генотипа.Как признал философ биологии Гайон: «Именно благодаря контакту с новой наукой о наследственности теория отбора стала по-настоящему понятной» (Gayon, 1992, p. 253). Объединение дарвиновского отбора с наследственностью известно как современный синтез в биологии (Huxley, 1942). Физическая основа генетической изменчивости включает мутации (изменения аллелей, вызванные экзогенными агентами, такими как химические вещества и радиация), рекомбинацию (смешение аллелей во время воспроизводства), генетический дрейф (случайный выбор аллелей, особенно на молекулярном уровне) и поток генов. (миграция генов между популяциями).Конечно, Дарвин ничего не знал о генетической основе наследования и не знал о работе Менделя, хотя одна неоткрытая книга со ссылкой на Менделя была найдена в его библиотеке (Zhang, p. 88). Даже после того, как широко распространились сведения о работе Менделя по наследованию фенотипических признаков, ген оставался лишь выводом из наблюдений (гипотетическая конструкция). По этой причине ген сначала не был принят в основную биологию никем иным, как Томасом Хантом Морганом, которого позже считали «отцом гена».
В современной интерпретации менделизма факты быстро трансформируются в факторы [то есть гены]. Если один фактор не объясняет факты, используются два; если двух окажется недостаточно, иногда срабатывают три. Превосходная фокусировка, иногда необходимая для объяснения результатов, может ослепить нас. . . до банальности того, что результаты так превосходно «объяснены», потому что объяснение было изобретено для их объяснения. Мы работаем в обратном направлении от фактов к факторам, а затем — готово! Объясняйте факты с помощью тех самых факторов, которые мы изобрели для их объяснения.(Морган, 1909; цитируется в Shine & Wrobel, 1976, стр. 51)
Поведенческие аналитики узнают в ранних оговорках Моргана по поводу родственных ему оговорок гена Скиннера о фэнтезийной физиологии того времени (Skinner 1938; 1950). И только после того, как экспериментальная работа Моргана позволила прямое наблюдение за гигантскими хромосомами слюнной железы плодовой мушки, он приветствовал ген в науке о наследственности.
Вариация в поведенческом анализе
С самого начала Скиннер (1935) считал вариацию фундаментальной.Мало что было известно о его биологической основе, но Скиннер рассматривал как среду, так и поведение как классов событий. Соединение экземпляров экологических и поведенческих событий с выбранными вариантами поощрения внутри этих классов. Состав классов менялся от момента к моменту. Таким образом, в последовательные моменты свет, который функционировал как различительный стимул в оперантной камере, мог быть замечен крысой под разными углами с несколько разной интенсивностью, и нажатие на рычаг, за которым следовало подкрепление, могло происходить с разными силами и топографией.Не только Скиннер признавал неизбежность изменения управляющих стимулов. Бывший ученик Скиннера Уильям Эстес позже разработал математическую теорию, которая явно реализовывала вариацию стимула, а именно теорию выборки стимула (Estes, 1950). (См. Guthrie, 1935, где дан молекулярный взгляд на природу реакции в его различении между движениями и действиями.) Более поздняя работа на нейронном уровне подробно документирует вариации внутри классов стимулов и реакций. То есть стимул с постоянными физическими характеристиками активирует переменную подгруппу рецепторов по мере изменения их пороговых значений и скорости привыкания.Точно так же одна и та же поведенческая реакция является согласованным продуктом активности различных членов популяции мотонейронов и мышечных волокон, которые они иннервируют (Geogopoulos, Schwartz, & Ketner, 1986). Наконец, нейроны головного и спинного мозга, которые взаимодействуют между сенсорными и двигательными нейронами, представляют собой переменную подгруппу клеток центральной нервной системы. Таким образом, на нейронном уровне последовательные экземпляры «одного и того же» стимула и реакции никогда не идентичны.
Вариация поведенческих реакций в данной среде (поведенческий фенотип) — это вариация, от которой зависят подкрепления.Именно из этой вариации подкрепители лишь косвенно выбирают нервно-мышечные события, с которыми коррелируют события окружающей среды и поведенческие события. По мере того, как эволюционная и обусловливающая истории организма становятся более обширными, различные стимулы, которые могут ощущаться в этой среде, начинают контролировать разные реакции с разной силой. Сила этих ответов зависит от того, какие стимулы воспринимаются в данный момент, и подробностей их истории по отношению к этим стимулам.Некоторые отношения между окружающей средой и поведением относительно постоянны в течение эволюционного времени и становятся рефлексивными отношениями в результате естественного отбора. Рефлексы могут быть относительно простыми, например, реакция зрачков на изменения освещения, или могут быть довольно сложными, например, рефлекс выпрямления всего тела в ответ на гравитационные сигналы. Однако основной источник вариаций возникновения сложного поведения следует искать в истории подкрепления. Например, если разные реакции были обусловлены разными элементами одной и той же среды, какая реакция возникает в данный момент, зависит от таких переменных, как предшествующая частота подкрепления для этих ответов.В зависимости от деталей этой истории один ответ может быть доминирующим в один момент, а другой — в другой, даже в одной и той же общей среде (например, Blough, 1966; Notterman, 1959; Palmer, в печати). История подкрепления организма становится все более важным фактором поведенческой изменчивости, так же как история естественного отбора вида становится все более важным фактором морфологической изменчивости.
Выбор
Выбор — это второй этап процесса отбора.В случае эволюционной биологии естественный отбор воздействует на фенотипические варианты и, менее прямо, на генетические варианты, с которыми они коррелируют. Естественный отбор изменяет относительные частоты генов в популяции различных организмов в следующем поколении. В случае поведения отбор путем подкрепления изменяет относительные частоты поведения (и косвенно физиологические события, с которыми они коррелируют) в поведенческом репертуаре того же самого организма в последующих случаях.Следующее обсуждение фокусируется на том, как действует отбор в этих двух случаях.
Естественный отбор
Агенты отбора в естественном отборе возникают из среды, в которой возникают варианты. Это включает не только внешнюю среду — как ее физические, так и органические элементы — но также и внутриорганическую среду организма, поскольку эти среды влияют на репродуктивную пригодность. Из-за различий в сочетании фенотипических признаков, сосуществующих в данном организме, а также из-за капризов среды отбора, результат отбора является вероятностным.Например, Джордж Вашингтон, несомненно, обладал многими качествами, которые обычно способствуют репродуктивному успеху, но он не оставил детей, хотя известно, что его жена была фертильной, учитывая, что у нее были дети от предыдущего брака. Результат естественного отбора еще больше осложняется тем фактом, что выживают (то есть сохраняются) и передаются следующему поколению не черты, а только гены. Более того, на большинство черт, особенно поведенческих, влияют несколько генов.Образно говоря, черты — это тени на стенах пещеры Платона, а гены — это объекты, отбрасывающие тени. Среда выбора «видит» только тени, но одни и те же объекты могут иногда отбрасывать разные тени. Феномен генетического автостопа прекрасно иллюстрирует эти осложнения (Wakely, p. 127). Предположим, что предпочтительный фенотип коррелирует с геном в определенном локусе хромосомы, а другой ген, расположенный в соседнем положении на той же хромосоме, не коррелирует таким образом.Поскольку два гена смежны, естественный отбор, вероятно, увеличит частоту обоих генов в последующих поколениях, потому что рекомбинация с меньшей вероятностью разделит их. «Отбор» полезного гена приводит к «отбору» гена автостопа, если использовать терминологию Собера (1984). Генетические исследования на людях показывают, что автостоп затрагивает около 10% генов в геноме.
Дарвинизм столкнулся с несколькими серьезными проблемами за последние 150 лет. Один из них — альтруизм, при котором организм действует, чтобы уменьшить свою индивидуальную репродуктивную пригодность, но при этом улучшает приспособленность других.Первой реакцией некоторых эволюционных биологов на это явление было продвижение молярной концепции естественного отбора. Если бы приспособленность группы , к которой принадлежал жертвующий человек, была увеличена, то, возможно, общая приспособленность могла бы быть улучшена (Wynne-Edwards, 1962). Однако Гамильтон (1964) показал, что эти явления не требуют перехода к более молярному, групповому взгляду на отбор. Вместо этого альтруизм можно понять, признав, что были сохранены гены как таковые, и что индивидуум, в котором они проживают, не имеет значения.Если у жертвователя и бенефициара общие гены, то альтруистическое поведение не представляет фундаментальной проблемы для естественного отбора. Обобщая свой краткий обзор альтруизма, Футуйма (стр. 18) пришел к выводу, что группы в молярных моделях «почти всегда являются родственными группами [т. Е. Генетически связанными], так что родственный отбор и групповой отбор — это два способа описания одного и того же процесса».
Второй вызов дарвиновскому отбору исходил от тех, кто сомневался, был ли эволюционный процесс постепенным, как предлагал Дарвин, или же он протекал дискретными шагами, ограниченными видоспецифическими характеристиками.Сторонники градуалистской позиции называли сальтационистскую позицию (то есть прерывистое равновесие) «эволюцией рывками» (Turner, 1984), на что сальтационисты ответили тем же, назвав градуалистов «эволюцией ползучим» (Gould & Rose, 2007). , стр.6). (Такова бесстрастность науки!). Подводя итоги этой дискуссии, Футуйма пришел к выводу, что в отношении требований сальтационистов о выборе на уровне видов «вызов синтетической теории провалился» (стр.19).
Последний серьезный вызов дарвиновскому объяснению биологической эволюции исходил от тех, кто доказывал важность для эволюции случайных генетических изменений на молекулярном уровне, так называемой теории нейтральных генов (Kimura, 1968). Все соглашались, что случайные генетические изменения способствуют изменчивости, но резко расходились во мнениях относительно того, как они влияют на ход эволюции. Историк науки Уильям Провайн восхищался этим спором: «Это величайшая тема в мире.Я даже не могу сказать, насколько это весело. Люди просто дерутся как сумасшедшие »(цитируется по Blum, 1992). В связи с этим спором Уэйкли (стр. 143) пришел к выводу, что от теории эволюции нейтральных генов следует отказаться, «учитывая недостаток силы теоретических аргументов в пользу нейтральной теории, эмпирических свидетельств против нее и того факта, что селективные модели могут … Лучше соответствуют наблюдениям и имитируют сам нейтралитет ». В качестве примера, один анализ показал, что «около 50% [молекулярных] замен будет вызвано положительным отбором у дрозофилы» (Wakeley, p.128).
Общий вывод из этих трех вызовов роли естественного отбора в эволюции состоит в том, что теория отбора возобладала, независимо от того, оспаривается ли она с более молярной или более молекулярной точки зрения. Как бы то ни было, столкновение с этими проблемами укрепило дарвиновскую точку зрения, показав, что ее можно расширить, чтобы охватить все более широкий спектр явлений на многих уровнях наблюдения (но см. Crow, 2008).
Выбор по армированию
Как происходит выбор при выборе по армированию? Скиннер считал, что «среда выбирает поведение» (Скиннер, 1985, стр.291), но он ничего не сказал о природе отборочного события. Он считал, что нейробиология в конечном итоге определит физическую основу отбора по подкреплению, но что большую часть поведения можно понять без характеристики подкрепляющего события. Подкрепления были просто стимулами, которые в последующих случаях увеличивали относительную частоту тех реакций, с которыми они были обусловлены. (См. Критику этой точки зрения в Meehl, 1950.) Позиция Скиннера имела множество прецедентов. Дарвин показал, что важное понимание эволюции может быть достигнуто путем принятия селекционной позиции, даже когда механизмы отбора были неизвестны.Скиннер считал, что ничто из того, что в конечном итоге было изучено о физиологии поведения, не может подорвать действующие поведенческие законы (Скиннер, 1938, стр. 432). Единственное необходимое условие для отбора по подкреплению, которое идентифицировал Скиннер (и многие другие), — это то, что подкрепление должно произойти сразу после поведения, которое нужно выбрать. «Сказать, что подкрепление зависит от реакции, может означать не что иное, как то, что оно следует за реакцией … обусловливание происходит только из-за временного отношения, выраженного в терминах порядка и близости реакции и подкрепления (Скиннер, 1948, стр. .168). Это обычно известно как требование временной непрерывности.
В конце 1960-х стало ясно, что необходимо что-то помимо связи предполагаемого подкрепления с поведением (Kamin, 1968; 1969). Было обнаружено, что если реакция была обусловлена предшествующим стимулом и за той же реакцией позже последовало такое же подкрепление в той же среде, но ей предшествовал вновь введенный предшествующий стимул, то новый стимул не получил бы контроля над ответом ( Рескорла и Вагнер, 1972; Вом Заал и Дженкинс, 1970).Что еще было нужно? Последующие исследования показали, что стимул действует как подкрепление, если он не только происходит во временном соединении с реакцией, но и вызывает активность, которая иначе не происходила бы в этой среде (Donahoe, Burgos, & Palmer, 1993; Donahoe, Crowley, Millard, & Стикни, 1982). Это верно, независимо от того, было ли изменение активности, вызванное подкрепляющим стимулом, результатом предшествующего естественного отбора (Stickney & Donahoe, 1983) или предшествующего отбора путем подкрепления (Palmer, 1987), и наблюдалась ли активность на уровне наблюдения за поведением. или только на нейронном уровне (например,г., Tobler, Fiorillo & Schultz, 2005). С этой точки зрения стимул действует как подкрепление только тогда, когда он вызывает изменение в активности. Например, если нажатие на планку крысой сопровождается «неожиданной» пищей, то нажатие на планку и слюноотделение усиливаются в этой среде. Естественный отбор создал механизм обучения, с помощью которого организм адаптируется к новым поведенческим требованиям окружающей его среды. И в той мере, в какой механизм обучения обеспечивает адаптацию, организм защищен от эффектов естественного отбора для этих отношений между средой и поведением (см.Ричардс, 1987). Именно в этом смысле «обучение является двигателем эволюции» (Wilson, 1975, стр. 156). Как и в случае с естественным отбором, отбор путем подкрепления «подготавливает» учащегося к тому, чтобы вести себя в будущем, как и в прошлом, в отношении случайных обстоятельств выбора.
Также, как и в случае с естественным отбором, «выбор» для определенного ответа может привести к «выбору» другого ответа, с которым он коррелирован. Это феномен суеверной обусловленности (Скиннер, 1948), аналог генетического автостопа при естественном отборе.Суеверное обусловливание является еще одним свидетельством общей природы стимулов и реакций, на которые действует отбор путем подкрепления. Я вспоминаю ситуацию, когда коллега, обученный ассоциативному обучению, попросил своего научного сотрудника обусловить нажатие на рычаг с несколькими крысами в оперантных камерах в моей лаборатории, но не использовать формирование. Таким образом мог произойти «естественный процесс обучения». Через некоторое время научный сотрудник пожаловался, что одна из крыс давила намного медленнее, чем другие, и, кроме того, из камеры периодически раздавались громкие «удары».Когда я заглянул в камеру через увеличительный глазок, я обнаружил, что крыса бросилась на прозрачный потолок камеры и вытянула ногу, чтобы поймать себя при падении. При этом крыса нажала на рычаг. Судя по всему, ранее обученная кормушка крыса пыталась выпрыгнуть из камеры и при этом случайно нажала на рычаг.
Приведенный выше взгляд на отбор по подкреплению основан на мгновенных случайностях окружающих и поведенческих событий с подкреплениями и согласуется с мнением Скиннера (1948) в этом отношении.Однако, как и в случае с вызовами группового отбора для дарвиновского отбора, основанного на группах и индивидуумах, концептуально аналогичные проблемы возникли в моментальных описаниях отбора путем подкрепления, основанного на группах ответов. Например, были обнаружены упорядоченные отношения между количеством подкреплений и количеством ответов во время воздействия параллельных графиков, так называемое отношение соответствия (Baum, 1973; 2002). Исследования выявили сиюминутные связи между стимулами, реакциями и подкреплениями, которые могут создавать молярные отношения (например,г., Бургос и Мурильо-Родригес, 2007; Кроули и Донахью, 2004; Хинсон и Стаддон, 1983; Макдауэлл, 2004). Однако вопрос о наиболее эффективном уровне, на котором можно сформулировать отбор путем подкрепления, остается спорным (Baum & Davison, 2009).
Удержание
Третья фаза процесса выбора — удержание. Сохранение позволяет выбранным вариантам сохраняться достаточно долго, чтобы они могли внести свой вклад в пул вариаций, на который будут действовать будущие выборы.
Сохранение в эволюционной биологии
Как уже отмечалось, при естественном отборе сохраняются гены, а не черты характера, которые представляют собой сложное выражение взаимодействия генов с окружающей средой индивида.Дарвиновское предвидение многих эволюционных явлений свидетельствует о силе селекционистского мышления, даже когда механизмы, реализующие его, неизвестны. История перехода Дарвина от его теории «геммул», согласно которой гипотетические сущности мигрировали из соматических клеток, чтобы влиять на половые клетки, к его версии наследования приобретенных характеристик, в результате чего он попытался опровергнуть математические аргументы Флиминга Дженкина (Gayon, p. 85 и далее), здесь не рассматриваются. Повторное открытие работы Менделя — после первоначального чрезмерного акцента на роли мутации со стороны ДеВри — привело к современной науке генетике, в которой гены передаются неизменными, за исключением мутаций, от родителей к потомкам.Генетика и молекулярная биология выявили объекты, тени которых отбрасываются на стены пещеры Платона.
Есть одно исключение из неизменяемости (бессмертия?) Генов, которое заслуживает здесь внимания — эпигенетика (Футуйма, стр. 21; Рэй, стр. 235; Ричерсон и Бойд, стр. 582). Эпигенетика изучает влияние родительской среды на гены, которые передаются их потомству. Было обнаружено, что эпигенетические изменения сохраняются на протяжении нескольких поколений (Seong et al, 2011).В качестве одного из примеров было обнаружено, что у крысят, которые подвергались стрессу при воспитании невнимательными матерями, были метильные группы (-Ch4), связанные с их генами. (Сами гены не пострадали). Позже метилированные гены были обнаружены и у их потомков (Gräff, Franklin, & Mansuy, 2010). Сообщалось о подобных эпигенетических эффектах у людей. Стресс метилирует материнские гены, и гены их потомства, в свою очередь, метилируются. Как следствие, реакция рецепторов кортикотропина у потомства изменяется, что влияет на их реакцию на стресс (Oberlander et al., 2008). Возможная эволюционная интерпретация этих эпигенетических эффектов заключается в том, что если какое-либо состояние присутствует в родительской среде, то такое же состояние, вероятно, также может возникнуть в среде потомства. Эпигенетика в настоящее время является активной и несколько спорной областью исследований, значение которой для различных явлений, включая память, изучается (например, Miller, 2010).
Удержание при выборе подкреплением
Сохранение эффектов отбора подкреплением зависит от стимулов в настоящей среде, а также от общественного и частного поведения, выбранного подкрепителями в прошлом в этой среде.(См. Обсуждение напоминания и запоминания в Donahoe & Palmer, 1994/2010.) Подобно тому, как наводнение может выявить окаменелости, ранее покрытые отложениями на берегу реки, так и исчезновение доминирующей реакции может выявить поведение, которое было слишком слабым, чтобы проявиться до тех пор, пока более сильные ответы ослабли (Cançado & Lattal, 2011; Epstein, 1985; Palmer, в печати; Shahan & Sweeney, 2011). Сохранение поведения опосредуется воздействием на нервную систему предшествующего отбора путем подкрепления. Если проследить путь от генов к признакам — непростая задача, то тем более путь от нейронов к поведению! В геноме человека «всего» около 20 000 структурных генов (хотя и много дополнительных регуляторных генов), но в человеческом мозге, возможно, имеется 100 миллиардов нейронов, связанных друг с другом более чем 100 триллионами синапсов.Следующий и последний раздел обзора описывает, как эволюция посредством естественного отбора столкнулась с проблемой отслеживания траектории эволюционных изменений, и обрисовывает в общих чертах аналогичную стратегию анализа поведения в попытке понять сложное поведение как продукт отбора путем подкрепления.
СЕЛЕКЦИОНИСТСКИЙ ПОДХОД К СЛОЖНЫМ ЯВЛЕНИЯМ
Что больше всего приветствовало поведенческого аналитика, читающего ESD , так это безоговорочное признание селекционизма — в форме естественного отбора — как источника сложности.О, этот отбор путем подкрепления занял бы такое высокое положение! Рассмотрим некоторые из следующих утверждений из ESD : «Вся молекулярная биология и геномика триумфально подтверждают единство жизни и ее общее происхождение» (Футуйма, стр. 7). «Естественный отбор является основной движущей силой эволюционных изменений… Несмотря на огромное разнообразие жизни на уровне организма и генома, существуют универсальные правила эволюции… [и]… эти правила могут быть обнаружены путем изучения относительно небольшого числа видов» ( Чжан, стр.88, 93). Сравните последнее утверждение с критикой Саймона (1980) о том, что бихевиоризм «озабочен лабораторными крысами, а не людьми, занятыми сложным мышлением и задачами решения проблем» (стр. 76). И, что наиболее ярко, сопоставьте взгляды Саймона со следующим комментарием Дарвина: «Тот, кто понимает бабуина, сделает для метафизики больше, чем Локк» (Richerson & Boyd, стр. 562, цитируя записные книжки Дарвина M и N в Gruber & Barrett, 1974, стр.281). Наконец, «… полное понимание любого вопроса в биологии требует объяснения его эволюционного происхождения…» (Zhang, p.108). Поскольку естественный отбор общепринят в эволюционной биологии, ESD не вынужден защищать его с помощью экспериментальной работы, но ему разрешено следовать его последствиям. Короче говоря, большая часть ESD посвящена научной интерпретации , а — не экспериментальному анализу, если использовать термины Скиннера для различия (Skinner, 1957).
Как естественный отбор занял центральное место в эволюционной биологии, в то время как селекционизм борется в науке о поведении? Наше понимание деталей отбора путем подкрепления, возможно, не менее полно, чем Дарвиновское понимание естественного отбора.Ответ лежит в двух направлениях. Во-первых, знание биологических механизмов, реализующих естественный отбор — генетики и молекулярной биологии — намного более продвинуто, чем наше нынешнее понимание нейронных механизмов отбора путем подкрепления. Во-вторых, количественные методы исследования последствий естественного отбора гораздо более развиты, чем методы отслеживания эффектов отбора путем подкрепления. Позвольте мне кратко прокомментировать каждый из них.
Стремясь создать независимую науку о поведении, некоторые пытались отделить анализ поведения от других биологических наук из-за опасений, что независимость анализа поведения каким-либо образом будет поставлена под угрозу (например,г., Риз, 1996; ср., Донахью, 1996). Скиннер считал анализ поведения «… строгим, обширным и быстро развивающимся разделом биологии…» (1974, стр. 255), но, несмотря на это, некоторые современные теоретики очерняют озабоченность биологической основой поведения: «Нейрофизиология может опускаться… потому что раскрывает только механизм »(Баум, 2011, с. 119). Хотя это правда, что экспериментальный анализ нейронных механизмов обусловливания представляет собой огромные проблемы, в этом направлении уже достигнут большой прогресс (например,г., Frey, 1997; Редондо, Окуно, Спунер, Френгуелли и Моррис, 2010 г .; Sweatt, 2010). Как бы то ни было, тем не менее верно также и то, что сложность нервной системы и наше непоправимое незнание полной истории отбора путем подкрепления навсегда исключают полный экспериментальный анализ любого конкретного случая сложного поведения. Эти трудности присущи не только анализу поведения, но и всем историческим наукам (Donahoe, 1997a; Donahoe & Palmer, 1989), включая эволюционную биологию.Однако, в отличие от эволюционной биологии, поведение редко оставляет какие-либо окаменелости, видимые невооруженным глазом (Tinbergen, 1963). Физические окаменелости поведения навсегда скрыты среди бесчисленных синапсов нервной системы.
Как эволюционная биология решает проблемы, связанные с концепцией? Эволюционная биология разработала количественные процедуры для отслеживания воздействия естественного отбора на популяцию вариантов. Действительно, разработка статистических процедур для отслеживания потока генов сыграла важную роль в современном синтезе генетики с естественным отбором (Fisher, 1930; Haldane, 1932; Wright, 1968).Эти математические методы позволили строго проследить за потоком генов между видами и поколениями и тем самым продемонстрировали, что изменения, предложенные Дарвином, на самом деле могут произойти. «Сегодня эволюционная теория правильно сформулирована в математических терминах и со ссылкой на генетику популяций» (Gardner, 2010). Благотворная роль количественных процедур в эволюционной биологии признана во многих местах в ESD , и не только в отношении генетики.«Палеонтологи воспользовались достижениями биологической теории и количественных методов, чтобы предсказуемо и количественно обобщить закономерности, которые Дарвин мог описать только словесно, и сформулировать предсказания для моделей Дарвина, которые сам Дарвин не смог получить» (Вагнер, стр. 451). «Хотя Чарльз Дарвин обладал удивительным видением, он не мог представить себе технологию, которая позволила бы биологам реконструировать и использовать Древо Жизни…» (Хиллис, стр. 445). Среди количественных методов, разработанных в ответ на сложность эволюционной биологии, есть исследования с использованием моделирования, которые стали возможными благодаря современным компьютерным технологиям (например,г., Lewontin & White, 1960; Мейнард-Смит, 1964). Компьютерное моделирование позволяет применять принципы, когда они слишком сложны для аналитических решений в замкнутой форме. Хотя моделирование может лишь частично уловить характерные особенности истории взаимодействия организмов с окружающей их средой, они предоставляют своего рода «доказательство существования» того, что сложные явления находятся в компетенции естественного отбора (Kokkop & Jennions, стр. 293; Wagner, с.455).
Анализ поведения также признал необходимость динамических количественных процедур (например,g., Marr, 1992) и начал использовать их для интерпретации результатов отбора путем подкрепления. Эти количественные процедуры были реализованы на различных уровнях анализа. Вся такая работа ограничивается поведенческими исследованиями, но она также может быть основана на результатах других уровней анализа. Некоторые количественные исследования в интерпретации поведения основываются на нейробиологии (например, Donahoe, 1997b), некоторые — на эволюционных соображениях, а также на нейробиологии, например, в гибридных генетических / обучающих алгоритмах (Burgos, 1997; Donahoe, 2002), некоторые — на эволюционных соображениях. с использованием генетических алгоритмов (McDowell, 2004) и других, в основном, исключительно из поведенческих соображений (Hutchison, 1997; Killeen & Fetterman, 1988).Количественная работа не обязательно должна отражать всю потенциально актуальную науку. Однако, чтобы квалифицировать как биоповеденческую интерпретацию — а не просто математическое моделирование — методы не должны явно противоречить такой информации. К чисто концептуальным ограничениям, например, что количественная процедура должна давать оптимальные решения, следует относиться с осторожностью, хотя они могут служить полезными ориентирами для сравнения количественных результатов (Kokkop & Jennions, p.299). Один биолог-эволюционист заметил, что убеждение в том, что «отбор порождает оптимальные решения, нельзя отстаивать как точное представление о реальном мире» (Gardner, 2010), и то же самое можно сказать и о поведенческом отборе. Организм является средоточием множества случайных обстоятельств на протяжении его индивидуальной и эволюционной истории, и вероятность того, что производительность будет оптимальной в любом одном случае, мала. Наконец, не существует единого «правильного» уровня, на котором можно было бы проследить эффекты отбора по подкреплению.Как Скиннер (1935) признал гораздо раньше, подходящим уровнем является тот, на котором возникает порядок в отношении изучаемого явления. Это относится как к количественным, так и к экспериментальным методикам.
Наконец, поскольку биологи-эволюционисты и поведенческие аналитики разделяют общую приверженность селекционизму, следует стремиться к плодотворному сотрудничеству, чтобы разгадать секреты обусловленности и памяти, а также то, как на них влияет генетика. Разработка препаратов оперантного кондиционирования для рыбок данио может показаться особенно многообещающей, потому что существует значительный объем соответствующей исследовательской литературы — по кондиционированию с рыбой (например.g., Hollis, 1997), о генетике рыбок данио (Detrich, Westerfield, & Zon, 2004), о связи генетики рыбок данио с поведением (например, Wolman & Granato, 2011) и о рецепторе NMDA, который известен играть центральную роль в кондиционировании позвоночных (Sison & Gerlai, 2011).
ССЫЛКИ
- Baum W.B. Корреляционный закон эффекта. Журнал экспериментального анализа поведения. 1973; 20: 137–153. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Baum W.Б. От молекулярного к молярному: смена парадигмы в поведенческом анализе. Журнал экспериментального анализа поведения. 2002. 78: 95–116. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Баум В. Что такое радикальный бихевиоризм ?: обзор книги Джея Мура «Концептуальные основы радикального бихевиоризма» . Журнал экспериментального анализа поведения. 2011; 95: 119–126. [Google Scholar]
- Баум В.М., Дэвисон М. Моделирование динамики выбора. Поведенческие процессы. 2009. 81: 189–194.[PubMed] [Google Scholar]
- Blough D.S. Стимулирование наименее частых интервалов между ответами. Журнал экспериментального анализа поведения. 1966; 9: 581–591. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Блюм Д. Ученые ведут открытую войну за «нейтральную теорию» генетики. Сакраменто пчела. 1992. 16 марта.
- Боулер П.Дж. Затмение дарвинизма. Издательство Университета Джона Хопкинса; Балтимор: 1983. [Google Scholar]
- Бургос Дж. Э. Развитие искусственных нейронных сетей в условиях Павлова.В: Донахью Дж. У., Дорсел В. П., редакторы. Нейросетевые модели познания. Elsevier Science Press; Нью-Йорк: 1997. С. 58–79. [Google Scholar]
- Бургос Дж. Э., Мурильо-Родригес Э. Нейросетевое моделирование двух контекстно-зависимых явлений. Поведенческие процессы. 2007. 75: 242–249. [PubMed] [Google Scholar]
- Кэмпбелл Д. Т. Эволюционная эпистемология. В: Шлипп П.А., редактор. Философия Карла Поппера (Том 14, стр. 413–463) Открытый суд; LaSalle, IL: 1974. [Google Scholar]
- Cançado C.R.X, Lattal K.A. Возрождение временных паттернов реагирования. Журнал экспериментального анализа поведения. 2011; 95: 271–287. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Карлсон К.Дж., Стаут Д., Джашашвили Т., де Руйтер Д.Дж., Таффоро П., Карлсон К., Бергер Л.Р. Эндокаст Mh2, Australopithecus sediba . Наука. 2011; 333: 1402–1407. [PubMed] [Google Scholar]
- Катания А.С. Некоторые дарвиновские уроки для анализа поведения: обзор книги Боулера Затмение дарвинизма .Журнал экспериментального анализа поведения. 1987. 47: 249–257. [Google Scholar]
- Катания А.С. Отбор по поведению и биологии. В: Тодд Дж. Т., Моррис Э. К., редакторы. Современный взгляд на Б. Ф. Скиннера и современный бихевиоризм. Greenwood Press; Вестпорт, Коннектикут: 1995. С. 185–194. (стр. 185–194). [Google Scholar]
- Кроу Дж. Ф. Споры середины века в популяционной генетике. Ежегодный обзор генетики. 2008; 42: 1–16. [PubMed] [Google Scholar]
- Кроули М.А., Донахью Дж.W. Matching: его приобретение и обобщение. Журнал экспериментального анализа поведения. 2004. 82: 143–159. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Деннетт Д.К. Опасная идея Дарвина: эволюция и смыслы жизни. Саймон и Шустер; Нью-Йорк: 1995. [Google Scholar]
- Детрих Х.В., Вестерфилд М., Зон Л.Л. Рыбки данио: генетика, геномика и информатика. Эльзевир; Бостон: 2004. [Google Scholar]
- Donahoe J.W. Правдоподобная аналогия. 1983 г.Теория подкрепления: когнитивная психология :: Естественный отбор: особенное творение. Приглашенный адрес представлен Американской психологической ассоциации, Анахайм, Калифорния, август.
- Донахью Дж. У. О связи поведенческого анализа и биологии. Поведенческий аналитик. 1996; 19: 71–73. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Donahoe J.W. Необходимость нейронных сетей. В: Донахью Дж. У., Дорсел В. П., редакторы. Нейросетевые модели познания. Elsevier Science Press; Нью-Йорк: 1997a.С. 1–21. [Google Scholar]
- Донахью Дж. У. Сети выбора: моделирование пластичности посредством обучения с подкреплением. В: Донахью Дж. У., Дорсел В. П., редакторы. Нейросетевые модели познания. Elsevier Science Press; Нью-Йорк: 1997b. С. 336–357. [Google Scholar]
- Донахью Дж. У. Анализ поведения и нейробиология. Поведенческие процессы. 2002; 57: 241–259. [PubMed] [Google Scholar]
- Донахью Дж. У. Селекционизм. В: Латтал К.А., Чейз П.Н., редакторы. Теория поведения и философия.Kluwer Academic Publishers; Дордрехт, Нидерланды: 2003. С. 103–128. (стр. 103–128). [Google Scholar]
- Донахью Дж. У., Бургос Дж. Э., Палмер Д. К. Селекционный подход к армированию. Журнал экспериментального анализа поведения. 1993; 58: 17-40. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Донахью Дж. У., Кроули М. А., Миллард В. Дж., Стикни К. А. Единый принцип армирования. В: Commons M.L, Herrnstein R.J, Rachlin H, editors. Количественный анализ поведения (т.2, с. 493–521) Баллинджер; Кембридж, Массачусетс, 1982. [Google Scholar]
- Донахью Дж. У., Палмер Д. К. Интерпретация сложного человеческого поведения: некоторые реакции на параллельную распределенную обработку , под редакцией Дж. Л. МакКлелланда, Д. Э. Румелхарта и PDP Group. Журнал экспериментального анализа поведения. 1989. 51: 399–416. [Google Scholar]
- Донахью Дж. У., Палмер Д. К. Обучение и сложное поведение. Аллин и Бэкон; Бостон: 1994/2010. (Третье переиздание, Ledgetop Publishing).[Google Scholar]
- Donahoe J.W, Wessells M.G. Обучение, язык и память. Харпер и Роу; New York: 1980. [Google Scholar]
- Edinborval V, Lefebvre J-F, Moreau C, Gbeha E, Hovhannesyan K, Bourgeois S,… Labuda D. Х-связанный гаплотип неандертальского происхождения присутствует среди всех неафриканских популяций. Молекулярная биология и эволюция. 2011; 7: 1957–1962. [PubMed] [Google Scholar]
- Эпштейн Р. Возрождение, вызванное исчезновением: предварительное исследование и возможное применение.Психологическая запись. 1985; 35: 143–153. [Google Scholar]
- Estes W.K. К статистической теории обучения. Психологический обзор. 1950; 57: 94–107. [Google Scholar]
- Фишер Р.А. Генетическая теория естественного отбора. Кларендон; Oxford: 1930. [Google Scholar]
- Frey U. Клеточные механизмы долгосрочной потенциации: позднее поддержание. В: Донахью Дж. У., Дорсел В. П., редакторы. Нейросетевые подходы к познанию: биоповеденческие основы. Elsevier Science Press; Амстердам: 1997.С. 105–128. (стр. 105–128). [Google Scholar]
- Гарднер А. Адаптация как дизайн организма. Письма о биологии. 2010; 5: 861–864. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Гайон, Жан. Борьба Дарвина за выживание. (Тр. Мэтью Кобб) Издательство Кембриджского университета; Кембридж, Великобритания: 1992. [Google Scholar]
- Геогопулос А.П., Шварц А.Б., Кетнер Р.Э. Нейронное популяционное кодирование движения. Наука. 1986; 233: 1416–1419. [PubMed] [Google Scholar]
- Гулд С.Дж., Роуз С., редакторы.Богатство жизни: главный Стивен Джей Гулд. Нортон; New York: 2007. [Google Scholar]
- Gräff J, Franklin T.B, Mansuy I.M. Эпигенетика и память. Biologie Aujourdhui. 2010; 2: 131–137. [PubMed] [Google Scholar]
- Gruber H.E, Barrett P.H. Дарвин о человеке: психологическое исследование творчества Дарвина вместе с ранними и неопубликованными записными книжками Дарвина. Даттон; Нью-Йорк: 1974. [Google Scholar]
- Гатри Э. Р. Психология обучения. Харпер и Роу; Нью-Йорк: 1935 год.[Google Scholar]
- Haldane J.B.S. Причины эволюции. Лонгман, Грин; Нью-Йорк: 1932. [Google Scholar]
- Hamilton W.F. Генетическая эволюция социального поведения, I и II. Теоретическая биология. 1964; 7: 1–52. [PubMed] [Google Scholar]
- Харфорд Т. Адапт: Почему успех всегда начинается с неудачи. Фаррар, Штраус и Жиру; Нью-Йорк: 2011. [Google Scholar]
- Hinson J.M, Staddon J.E.R. Соответствие, максимизация и восхождение на холмы. Журнал экспериментального анализа поведения.1983; 40: 321–331. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Holland J.H. Адаптация в естественных и искусственных системах. Пресса Мичиганского университета; Анн-Арбор, Мичиган: 1975. [Google Scholar]
- Holland J.H. Генетические алгоритмы. Компьютерные программы, которые развиваются способами, напоминающими естественный отбор, могут решать сложные проблемы, которые даже их создатели не полностью понимают. Scientific American, июль. 1992. С. 66–72.
- Холлис К.Л. Современные исследования Павловской обусловленности.Американский психолог. 1997. 52: 956–965. [PubMed] [Google Scholar]
- Халл Д.Л. Дарвин и его критики. Издательство Гарвардского университета; Кембридж, Массачусетс: 1973. [Google Scholar]
- Hutchison W.R. Нам также нужны полные поведенческие модели. Журнал экспериментального анализа поведения. 1997. 67: 224–228. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Huxley J.S. Эволюция: современный синтез. Аллен и Анвин; Лондон: 1942. [Google Scholar]
- Камин Л.Дж. «Подобные вниманию» процессы в классической обусловленности.В: Джонс М.Р., редактор. Симпозиум Майами по предсказанию поведения. Университет Майами Пресс; Корал-Гейблс, Флорида: 1968. С. 9–31. (стр. 9–31). [Google Scholar]
- Камин Л.Дж. Предсказуемость, удивление, внимание и обусловленность. В: Кэмпбелл Б.А., Чёрч Р.М., редакторы. Наказание и отвращение. Appleton-Century-Crofts; Нью-Йорк: 1969. С. 279–296. (стр. 279–296). [Google Scholar]
- Киллин П.Р., Феттерман Дж. Г. Поведенческая теория времени. Психологический обзор. 1988. 95: 274–295.[PubMed] [Google Scholar]
- Кимура М. Скорость эволюции на молекулярном уровне. Природа. 1968; 217: 624–626. [PubMed] [Google Scholar]
- King M.C., Wilson A.C. Двухуровневая эволюция человека и шимпанзе. Наука. 1975. 188: 107–116. [PubMed] [Google Scholar]
- Левонтин Р.К., Уайт М.Дж. Взаимодействие инверсионных полиморфизмов двух пар хромосом у кузнечика, Moraba scurra . Эволюция. 1960; 14: 116–129. [Google Scholar]
- МакДауэлл Дж.J. Вычислительная модель отбора по следствиям. Журнал экспериментального анализа поведения. 2004. 81: 292–317. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Марр М.Дж. Динамика поведения: одна точка зрения. Журнал экспериментального анализа поведения. 1992; 57: 249–266. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Мейнард-Смит Дж. Групповой отбор и родственный отбор. Природа. 1964; 200: 1145–1147. [Google Scholar]
- Майр Э. Виды животных и эволюция. Издательство Гарвардского университета; Кембридж, Массачусетс: 1963.[Google Scholar]
- Майр Э. Р. К новой философии биологии. Издательство Гарвардского университета; Cambridge, MA: 1988. [Google Scholar]
- Meehl P.E. О круговороте закона эффекта. Психологический бюллетень. 1950; 47: 52–75. [Google Scholar]
- Миллер Г. Роль эпигенетики в познании. Наука. 2010; 329: 27. [PubMed] [Google Scholar]
- Нунан Дж. П., Куп Дж., Кударавалли С., Смит Д., Краузе Дж., Алесси Дж.… Рубин Е. М. Секвенирование и анализ геномной ДНК неандертальцев.Наука. 2006; 314: 1113–1118. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Notterman J.M. Излучение силы при прессовании стержня. Журнал экспериментальной психологии. 1959; 58: 341–347. [PubMed] [Google Scholar]
- Оберландер Т.Ф., Вайнберг Дж., Папсдорф М., Грунау Р., Мисри С., Девлин А.М. Пренатальное воздействие материнской депрессии, неонатальное метилирование гена рецептора глюкокортикоидов человека (NR3C1) и стрессовые реакции детского кортизола. Эпигенетика. 2008; 3: 97–106. [PubMed] [Google Scholar]
- Палмер Д.C. Блокирование условного подкрепления. 1987. Неопубликованная докторская диссертация, Массачусетский университет, Амхерст.
- Палмер Д.К. Рассмотрение частных мероприятий требуется во всеобъемлющей науке о поведении. Поведенческий аналитик. под давлением. [Бесплатная статья PMC] [PubMed]
- Палмер Д.К., Донахью Дж. У. Эссенциализм и селекционизм в когнитивной науке и анализе поведения. Американский психолог. 1992; 47: 1344–1358. [PubMed] [Google Scholar]
- Редондо Р.Л., Окуно Х., Спунер П.A, Frenguelli B.G., Morris R.G.M. Синаптическое маркирование и захват: дифференциальная роль различных киназ кальция / кальмодулина в долгосрочном потенцировании, зависимом от синтеза белка. Журнал неврологии. 2010; 30: 4981–4989. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Reese H.W. Как физиология связана с анализом поведения. Поведенческий аналитик. 1996; 19: 61–70. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Рескорла Р.А., Вагнер А.Р. Теория Павловского кондиционирования: Вариации эффективности подкрепления и не подкрепления.В: Блэк А.Х., Прокасий В.Ф., ред. Классическое кондиционирование II. Appleton-Century-Crofts; Нью-Йорк: 1972. С. 64–99. (стр. 64–99). [Google Scholar]
- Richards R.J. Дарвин и появление эволюционных теорий разума и поведения. Издательство Чикагского университета; Чикаго: 1987. [Google Scholar]
- Сеонг К.Х., Ли Д., Шимицу Х., Накамура Р., Исии С. Наследование вызванных стрессом, зависимых от ATF-2 эпигенетических изменений. Клетка. 2011; 145: 1049–1061. [PubMed] [Google Scholar]
- Шахан Т.А, Суини М. Модель возрождения, основанная на поведенческом импульсе. Журнал экспериментального анализа поведения. 2011; 95: 91–108. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Shine I, Wrobel S. Томас Хант Морган: пионер генетики. Университет Кентукки Пресс; Lexington, KY: 1976. [Google Scholar]
- Сидман М. Тактика научных исследований. Базовые книги; Нью-Йорк: 1960. [Google Scholar]
- Саймон Х. Поведенческие и социальные науки. Наука. 1980; 209: 72–78.[PubMed] [Google Scholar]
- Sison M, Gerlai R. Эффективность ассоциативного обучения у рыбок данио ( Danio rerio ) ухудшается антагонистом NMDA-R MK-801. Нейробиология обучения и памяти. 2011; 2: 230–237. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Скиннер Б.Ф. Общая природа концепций стимула и реакции. Журнал общей психологии. 1935; 12: 40–65. [Google Scholar]
- Скиннер Б.Ф. Поведение организмов. Appleton-Century-Crofts; Нью-Йорк: 1938 год.[Google Scholar]
- Скиннер Б.Ф. «Суеверие» у голубя. Журнал экспериментальной психологии. 1948; 38: 168–172. [PubMed] [Google Scholar]
- Скиннер Б.Ф. Необходимы ли теории обучения? Психологический обзор. 1950; 57: 193–216. [PubMed] [Google Scholar]
- Скиннер Б.Ф. Наука и поведение человека. Макмиллан; Нью-Йорк: 1953. [Google Scholar]
- Скиннер Б.Ф. Вербальное поведение. Appleton-Century-Crofts; Нью-Йорк: 1957. [Google Scholar]
- Скиннер Б.F. Онтогенез и филогенез поведения. Наука. 1966; 153: 1203–1213. [Google Scholar]
- Скиннер Б.Ф. О бихевиоризме. Случайный дом; Нью-Йорк: 1974. [Google Scholar]
- Скиннер Б.Ф. Когнитивная наука и бихевиоризм. Журнал психологии. 1985; 76: 291–301. [PubMed] [Google Scholar]
- Смолин Л. Жизнь в космосе. Издательство Оксфордского университета; Нью-Йорк: 1997. [Google Scholar]
- Собер Э. Природа отбора: эволюционная теория в философском фокусе.MIT Press; Кембридж, Массачусетс: 1984. [Google Scholar]
- Стаддон Дж. Э. Р., Эттингер Р. Х. Обучение: Введение в принципы адаптивного поведения. Харкорт, Брейс, Йованович; New York: 1989. [Google Scholar]
- Stickney K.J, Donahoe J.W. Ослабление блокировки за счет изменения локуса США. Обучение и поведение животных. 1983; 11: 60–66. [Google Scholar]
- Sweatt J.D. Механизмы памяти (2-е изд.) Elsevier Academic Press; Сан-Диего, Калифорния: 2010. [Google Scholar]
- Tinbergen N.О целях и методах этологии. Zeitschrift für Tierpsychologie. 1963. 20: 410–433. [Google Scholar]
- Тоблер П.Н., Фиорилло К.Д., Шульц В. Адаптивное кодирование значения вознаграждения с помощью дофаминовых нейронов. Наука. 2005; 307: 1642–1645. [PubMed] [Google Scholar]
- Тернер Дж. Зачем нужна эволюция рывками. Новый ученый. 1984; 101: 34–35. [Google Scholar]
- Vom Saal W, Jenkins H.M. Блокирование развития стимульного контроля. Обучение и мотивация. 1970; 1: 52–64. [Google Scholar]
- Уилсон Э.О. Социобиология. Издательство Гарвардского университета; Кембридж, Массачусетс, 1975. [Google Scholar]
- Вулман М., Гранато М. Поведенческая генетика в обучении личинок рыбок данио. Возрастная нейробиология. 2011. doi: 10.1002 / dneu.20872. [Бесплатная статья PMC] [PubMed]
- Райт С. Эволюция и генетика популяций. Издательство Чикагского университета; Чикаго, Иллинойс: 1968. [Google Scholar]
- Винн-Эдвардс V.C. Рассеивание животных в зависимости от социального поведения. Оливер и Бойд; Лондон: 1962 год.[Google Scholar]
- Yotova V, Lefebvre J.F, Moreau C, Gbeha E, Hovhannesyan K, Bourgeois S,… Labuda D. Х-связанный гаплотип неандертальского происхождения присутствует во всех неафриканских популяциях. Молекулярная биология и эволюция. 2011; 28: 1957–1962. [PubMed] [Google Scholar]
Parapatric Speciation — обзор
6.4 Генетика водорослей и выбор штаммов
Имеется относительно немного опубликованных исследований, касающихся разведения ламинарии и генетического улучшения, большинство из которых сосредоточено на Saccharina japonica (для обзор см. в Robinson, Winberg, & Kirkendale, 2013).В Китае и соседних странах, где этот вид широко культивируется, основная стратегия заключалась в проведении повторяющихся поколений нескольких инбредных линий (Robinson et al., 2013). Это привело либо к улучшению производства биомассы, либо к увеличению конкретных свойств / молекул. На сегодняшний день знания о генетике ламинарии в Чили сосредоточены на эволюционных и экологических темах: (1) глобальная филогеография и парапатрическое видообразование; (2) региональная филогеография и последниковая реколонизация; и (3) местное генетическое разнообразие и структура после природных и антропогенных нарушений.Первые опубликованные исследования были направлены на понимание воздействия природных (например, ENSO, Martínez et al., 2003) и антропогенных нарушений (например, отходы добычи меди, Faugeron, Martínez, Correa, & Billot, 2005) на пространственную и временную динамику ламинария L. nigrescens на севере Чили. Оба исследования охарактеризовали генетическую структуру популяций внутри и между сплошными насаждениями, разделенными зонами воздействия. Они показали, что нарушение в остальном непрерывного распространения этой водоросли совпало с усилением генетической дифференциации между насаждениями, происходящими к северу и югу от пораженного района.Эти авторы продемонстрировали, что поток генов происходил в основном между популяциями, разделенными небольшими расстояниями (т.е. максимум несколько километров) в результате их относительно короткой способности к расселению. Действительно, после события на ЭНСО 1982/83 г., даже через 20 лет L. nigrescens не переселили более 50 км береговой линии. Это медленный темп, учитывая, что это событие привело к гибели обширных зарослей водорослей вдоль 600 км береговой линии, в результате чего в центрах апвеллинга остались лишь редкие реликтовые популяции (Martínez et al., 2003). Эти результаты были дополнительно подтверждены филогеографическими исследованиями, охватывающими большую часть распространения видов (Tellier, Meynard, Correa, Faugeron, & Valero, 2009). Основываясь на ядерных, митохондриальных и пластидных маркерах, в каждой выборке наблюдалось низкое генетическое разнообразие, которое было характерно только для соседних популяций. Кроме того, сильная генетическая дивергенция наблюдалась в биогеографической переходной зоне 30 ° ю.ш., Описанной Камю (2001) и Тиль и др. (2007). Это генетическое расхождение соответствовало процессу видообразования (Tellier, Tapia, Faugeron, Destombe, & Valero, 2011) в результате как уменьшенного расселения, так и экологического расхождения (López-Cristoffanini, Tellier, Otaiza, Correa, & Contreras-Porcia , 2013; Опплигер и др., 2012, 2011). В целом, эти результаты показали, что определенные признаки (в данном случае признаки, связанные с температурой и устойчивостью к высыханию) могут развиваться в ответ на давление естественного отбора в диких популяциях.
Генетические исследования M. pyrifera на побережье Чили были сосредоточены на глобальной филогеографии. Что касается L. nigrescens, Macaya и Zuccarello (2010) показали низкое локальное (т.е. внутрипопуляционное) генетическое разнообразие и сильную генетическую дифференциацию между популяциями M.pyrifera , и / или между регионами вдоль чилийского побережья к северу от 42 ° ю. предположили, что локальные, а не глобальные процессы формируют генетическое разнообразие преимущественно из-за ограниченного распространения. В глобальном масштабе снижение генетического разнообразия было подчеркнуто относительно недавним прибытием этого рода в южное полушарие в результате распространения транс — тропиков из северного полушария в течение позднего плейстоцена (см. Coyer, Smith, & Андерсен, 2001; Macaya & Zuccarello, 2010).Иначе обстоит дело к югу от 42 ° ю.ш., где постледниковая колонизация привела к присутствию единственного гаплотипа в районе Магелланова пролива (Macaya & Zuccarello, 2010). Здесь важность распространения на большие расстояния была определяющим фактором в формировании генетического разнообразия такого большого географического региона. Действительно, этот единственный гаплотип является общим с другими субантарктическими регионами, включая юг Новой Зеландии, что очень похоже на то, о котором сообщалось для водорослей быка, Durvillaea antarctica (Fraser, Nikula, Spencer, & Waters, 2009).Однако в устойчивых популяциях, для которых достигается равновесие между соответствующим влиянием потока генов, генетического дрейфа и мутации, казалось, что ограниченное распространение спор было доминирующим процессом, формирующим местное и региональное генетическое разнообразие (см. Alberto et al., 2010, 2011). Это было, по-видимому, обычным явлением у большинства видов водорослей (см. Valero et al., 2011).
Интересной особенностью филогеографических исследований было то, что они сообщили об общих гаплотипах между всеми известными морфотипами, которые ранее считались разными видами (например,г. Macrocystis integrifolia , Macrocystis angustifolia и Macrocystis laevis sensu Demes, Graham, & Suskiewicz, 2009), что предполагает их недавнего общего предка. Однако этот результат все еще обсуждается, как предположили Astorga, Hernández, Valenzuela, Avaria-Llautureo и Westermeier (2012), которые использовали ядерные маркеры (ITS-2), и представители Macrocystis из Канады, США, Южной Африки, Австралия и Чили. Авторы наблюдали сильный филогенетический сигнал морфологических различий, который указывает на то, что исторические процессы, связанные с диверсификацией, могли определять текущую дифференциацию морфотипа.Недавние свидетельства гетерозиса у гигантских водорослей (Westermeier et al., 2010, 2011) были дополнительным признаком того, что фенотипическая дифференциация существует между генетически дифференцированными популяциями в Чили и что местное разнообразие может быть переорганизовано путем контролируемых скрещиваний в новые генотипы, давая ценные фенотипы (Westermeier et al., 2011). Это сильно контрастировало с парадигмой высокой фенотипической пластичности, лежащей в основе разнообразия Macrocystis , как предположили Graham et al.(2007) и Demes et al. (2009). Помимо морфологической пластичности, в различных прибрежных местообитаниях наблюдались пластические фенотипические реакции с точки зрения физиологических стратегий и стратегий жизненного цикла (Buschmann et al., 2006, 2014, 2004). Следовательно, относительная роль пластичности и отбора в формировании фенотипов ламинарии все еще остается дискуссией, требующей особого научного внимания.
Будущие перспективы создания программы селекции Macrocystis должны включать подробный анализ фенотипической дифференциации (морфология, химический состав и другие соответствующие производственные признаки) между естественными популяциями вместе с оценкой генетического вклада (т.е. наследственность) к значениям черт. Важно продолжить исследования природных популяций, потому что процесс одомашнивания этого вида только начинается, и до настоящего времени большая часть производства водорослей была основана на спорах, полученных из естественных насаждений. С другой стороны, генетическое взаимодействие культивируемых и естественных запасов потребует внимания, чтобы избежать потенциальных генетических модификаций естественных запасов за счет потока генов от модифицированных сортов при выращивании на открытом воздухе, что также было предложено для таких водорослей, как G.chilensis (Guillemin et al., 2008). Наконец, управление жизненным циклом ламинарии — это проблема, требующая особого внимания из-за ее способности использовать партеногенез для производства чистых генетических линий (т.е. полностью гомозиготных генотипов) в одном поколении, как предполагают недавние исследования, показывающие, что партеногенетические спорофиты могут производить однополые ( например, 100% самок) гаметофитное потомство (Shan, Pang, & Gao, 2013). Чистые генетические линии необходимы для количественной генетики конкретных признаков (например,г. QTL-анализ) и определение генетических маркеров для помощи в селекционных стратегиях.
Репродуктивная изоляция — обзор
Следует ожидать разногласий относительно того, что именно представляет собой вид, учитывая, что эта концепция выполняет очень много функций (Vane-Wright, 1992). Нас может интересовать классификация как таковая или эволюционное значение видов; в теории видов или просто в том, как их распознать; или по их репродуктивному, физиологическому или хозяйственному статусу.
Большинство неспециалистов, вероятно, имеют смутное представление о том, что виды определяются тем, что они не скрещиваются друг с другом; обычно гибриды между разными видами бесплодны или вообще неспособны к гибридизации. Такое впечатление в конечном итоге происходит из определения Mayr (1940), согласно которому виды — это «группы фактически или потенциально скрещивающихся естественных популяций, которые репродуктивно изолированы от других таких групп» (Концепция биологических видов). На самом деле Майр никогда не говорил, что виды не могут скрещиваться между собой , более того, он отрицал, что это каким-либо образом является необходимой частью репродуктивной изоляции; он просто сказал, что в естественных условиях они не , а .
Аллопатрия
Сказать, что две популяции аллопатричны, означает, что их географическое распределение не пересекается — они полностью разделены. Это означает, что у них нет возможности спариваться друг с другом, даже если бы они захотели. Например, невозможно проверить, действительно ли Macaca fuscata (Япония), M.cyclopis (Тайвань) и M.mulatta (макака-резус, материковая часть Восточной Азии) являются разными видами. или нет; они классифицируются как отдельные виды во всех основных контрольных списках, но нет объективного способа проверить эту классификацию в рамках Концепции биологических видов.
Действительно, это обычная ситуация: популяции, которые в некотором отношении отличаются друг от друга и по соответствующим критериям близкородственны, обычно аллопатричны. Чтобы получить доказуемую репродуктивную изоляцию, необходимый критерий в соответствии с Концепцией биологических видов, как sine qua non статуса вида, будет заключаться в том, чтобы оставить большинство живых организмов неклассифицируемыми, за исключением некоторого произвольного распоряжения.
Естественное скрещивание
Два общих вида североамериканских оленей ( Odocoileus virginanus , Whitetail и O.hemionus , Blacktail) встречаются вместе на обширной географической территории и всегда легко различимы; тем не менее, молекулярные исследования обнаружили доказательства того, что гибридизация имела место. Например, в Кантри Пекос, западный Техас, четыре из девяти исследованных белохвостов имели митохондриальную ДНК, характерную для черных хвостов, с которыми они делят свой ареал (Carr and Hughes, 1993). Очевидно, в недалеком прошлом самки чернохвоста присоединились к племенным стадам белохвоста, и, хотя фенотип белохвоста был строго выбран, мтДНК чернохвоста осталась в популяции, что является ископаемым документом о событии гибридизации.
У приматов также есть примеры гибридизации в дикой природе. Хороший пример первого случая, Cercopithecus ascanius (Краснохвостая обезьяна) и C.mitis (Голубая обезьяна) в Уганде, был подробно описан Struhsaker et al. (1988). Две обезьяны, которые очень симпатичны, что означает, что они живут в одних и тех же областях в широком диапазоне, скрещиваются на довольно заметных уровнях, но остаются отдельными и четко различимыми, и никто никогда не предлагал рассматривать их как отдельные виды.Этот случай мало чем отличается от упомянутого выше североамериканского оленя.
Это два примера — один непримат, другой примат — пар различных видов, которым удается сохранять отличия на обширных территориях, даже если между ними существует поток генов. Гораздо более обычными (или, лучше, более легко задокументированными) являются случаи, когда пары видов занимают ареалы, которые в значительной степени разделены, но встречаются вдоль своих границ (парапатрические), и скрещиваются там, где это происходит. Скрещивание варьируется от случайных до полных гибридных зон, и такие случаи, в отличие от случаев гибридизации в симпатрии, рассматриваются как свидетельство того, что репродуктивной изоляции не существует, поэтому эти два вида следует объединить в один.Но принципиального отличия от случаев гибридизации в симпатрии нет.
Классическим исследованием гибридной зоны является исследование двух мышей, Mus musculus и Mus domesticus , на полуострове Ютландия, Дания (см. Резюме в Wilson et al. , 1985). Гибридная зона, судя по морфологии и аллелям белков, очень узкая; однако мтДНК южного вида, M.domesticus , хорошо проникает через границу и через морской путь (Скагеррак) в Швецию.Это говорит о том, что происходила гибридизация, и что M.musculus расширяли свой ареал, а гибридная зона двигалась на юг с тех пор, как в раннем голоцене море разделило Данию и Швецию. В течение этого длительного периода не проводилось отбора против гибридизации.
В хорошо изученном примере приматов два павиана, Papio hamadryas (Hamadryas baboon) и P.anubis (Olive baboon), парапатричны и гибридизуются там, где их ареалы встречаются в Эфиопии, причем гибридная зона составляет не более шириной несколько километров.Эти два таксона адаптированы к более засушливой и более мезической среде, соответственно, и гибридная зона перемещается вверх и вниз по реке Аваш в зависимости от того, была ли серия засушливых сезонов или серия влажных сезонов, но остается более или менее подходящей. такая же ширина. Таким образом, этот случай мало чем отличается от случая с двумя мышами в Дании. В отличие от примера Cercopithecus , два таксона павианов перемещались между подвидами и видами (сравните Jolly, 1993 и Groves, 2001). Но в чем разница на самом деле?
Исследование подтверждает, что армирование тканей GORE® BIO-A® является проверенной альтернативой биопрепаратам при герниопластике.
Flagstaff, Ariz.И Милан, Италия (28 апреля 2015 г.) — Компания W. L. Gore & Associates, Inc. (Gore) объявила сегодня окончательные результаты долгосрочного проспективного многоцентрового исследования, посвященного изучению качества жизни пациентов после лечения сложной вентральной грыжи (ВГ). Исследователи представили свои результаты в рамках исследования «Комплексная открытая биоабсорбируемая реконструкция брюшной стенки», посвященного пластике грыжи с помощью биосинтетической сетки, на Всемирной конференции по грыжам в 2015 году в Милане, Италия. Биосинтетическое армирование тканей GORE® BIO-A® было изучено при лечении грыж в девяти центрах в США и Европе.Пациенты сообщили об улучшении качества жизни с течением времени после комплексного восстановления VH с использованием GORE® BIO-A® Tissue Reinforcement.
Пациенты, которым запланировано комплексное восстановление ВГ, часто имеют низкое качество жизни перед операцией из-за фистулы желудочно-кишечного тракта, инфекции сетки или незаживающей раны брюшной полости. GORE® BIO-A® Tissue Reinforcement — это уникальный вариант материала для сложных случаев, поскольку он разработан с идеальной пористостью для быстрой клеточной инфильтрации и васкуляризации для быстрого создания качественной ткани.
GORE® BIO-A® Tissue Reinforcement — это ткань уникальной конструкции из биосовместимых синтетических полимеров, которые организм постепенно впитывает, а ее трехмерная матрица из открытых, тесно связанных между собой пор способствует образованию и заживлению тканей. Как синтетический тканевый каркас, он не получен из тканей человека или животных, а спроектирован так, чтобы обеспечить единообразие, стабильность и универсальность. Клинические данные демонстрируют, что каркас заменяется подходящим коллагеном I типа, не оставляя постоянного материала.GORE® BIO-A® Tissue Reinforcement в настоящее время продается во многих странах мира.
Наше исследование, возможно, является одним из первых комплексных клинических исследований трехмерной биосинтетической сетки VH, учитывающих удовлетворенность пациентов. Этот универсальный материал, имеющий трехлетний срок хранения и не требующий замачивания, охлаждения или отслеживания, является простой в использовании альтернативой с проверенными характеристиками, которая одинаково выгодна как для хирургов, так и для больниц.
«Это исследование представляет собой важный шаг вперед в определении наиболее подходящего способа восстановления сложных дефектов брюшной стенки», — сказал Майкл Розен, доктор медицинских наук, Клиника Кливленда.«GORE® BIO-A® Tissue Reinforcement очень хорошо зарекомендовал себя в этой сложной группе пациентов и станет экономически эффективной альтернативой для реконструктивных хирургов».
Биологические сетки для грыж имеют высокую стоимость из-за сбора тканей трупов, свиней, коров и последующей обработки для удаления клеток для защиты пациентов от возможного вирусного заражения. Большинство других синтетических материалов, хотя и менее дорогие, являются постоянным дополнением к телу пациента. Технология биорассасывающегося материала GORE® BIO-A® сочетает в себе лучшее из двух вариантов лечения грыжевой сетки, последовательно создаваемых без оставления синтетического материала и без риска вирусного заражения биологическими материалами тканевого происхождения.
«Эта революционная технология представляет новый класс биосинтеза, который одновременно обеспечивает высокую удовлетворенность пациентов и экономит больницы и плательщикам потенциально значительные суммы денег, которые они тратят на биопрепараты каждый год для комплексной пластики грыжи и реконструкции брюшной стенки», — сказал Рон Андерсон, генеральный директор Руководитель подразделения хирургических изделий Gore.
О НАС
Компания Gore более 35 лет предлагает творческие терапевтические решения сложных медицинских проблем.За это время было имплантировано более 35 миллионов инновационных медицинских устройств Gore, которые спасли и улучшили качество жизни во всем мире. В нашу обширную линейку продуктов входят сосудистые трансплантаты, эндоваскулярные и интервенционные устройства, хирургические сетки для реконструкции грыж и мягких тканей, материалы для укрепления штапельной линии и швы для использования в сосудистой, сердечной и общей хирургии. Мы — одна из немногих избранных компаний, вошедших во все списки «100 лучших компаний для работы» США с момента дебюта рейтинга в 1984 году.
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались.
Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie. - Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г.,
браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере. - Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.
Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie
потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт
не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к
остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
карточек видов и видов | Quizlet
Несмотря на то, что симпатрические популяции не изолированы географически, они могут стать репродуктивно изолированными, адаптируясь к различным экологическим нишам посредством деструктивного отбора.
Пример: яблочная личинка мухи полагается на яблоки для завершения своего жизненного цикла — яблоки являются важной частью их ниши. Мужские и женские личинки яблони обычно ухаживают за плодами яблони и спариваются с ними. Затем самка откладывает оплодотворенное яйцо внутри плода, которое будет источником пищи для растущей личинки. После того, как плод упадет с дерева, личинка зарывается в землю и окукливается.Следующей весной появляется новый взрослый особь, начиная цикл заново.
Филогенетические деревья, оцененные на основе синапоморфий в данных последовательности ДНК, показывают, что мухи-личинки яблони очень тесно связаны с мухами-личинками боярышника, которые обитают в Северной Америке. Эти данные предполагают, что яблочные личинки произошли от личинок боярышника после завоза яблок (яблони были завезены в Северную Америку 300 лет назад из Европы). Мухи боярышника откладывают яйца в плоды боярышника, а яблони откладывают яйца в яблоки, хотя боярышник и яблони растут почти бок о бок.
Только 6% наблюдавшихся спариваний происходят между личинками яблони и боярышника.
— Яблочные мухи сильнее всего реагируют на запахи яблок. Мухи боярышника сильнее всего реагируют на запахи боярышника.
— У обоих типов мух нет разницы в реакции на сочетание обоих запахов или вообще отсутствует запах.
— Яблочные мухи избегают запаха боярышника, а запах боярышника избегают запаха яблок.
— Способность мух различать запахи имеет генетическую основу — это означает, что яблоневые мухи и мухи боярышника имеют разные аллели, связанные с влечением к фруктам. фруктовые ароматы так же, как их родители.
В результате, хотя яблони и боярышник живут в одной и той же географической области, презиготическая репродуктивная изоляция происходит в результате естественного отбора для адаптации к двум разным нишам. Мухи яблони спариваются с яблоками (избегая плодов боярышника), а мухи боярышника спариваются с плодами боярышника (избегая плодов яблони).